Download

1 / 26
260 likes | 457 Views
Unidad I Fundamentos neurales de la percepción visual La retina y el LGN. 20x20’’ libre de conos S. 1º. Conos LMS en proporción 10:5:1; los conos L y M están distribuidos de manera aleatoria, los conos S, sin embargo, parecen dispuestos de manera semi-regular. 1º. Propiedades de...

E N D
Unidad I Fundamentos neurales de la percepción visual La retina y el LGN
20x20’’ libre de conos S 1º Conos LMS en proporción 10:5:1; los conos L y M están distribuidos de manera aleatoria, los conos S, sin embargo, parecen dispuestos de manera semi-regular. 1º Propiedades de... L.T. Sharpe et al. (1999). Fig. I.1b.
540 440 565 Propiedades de los fotorreceptores. Sensibilidad espectral de los conos: absortancias efectivas. A. Stockman y L.T. Sharpe (2000). Fig. I.2.
Principio de Univariancia Propiedades de los fotorreceptores. Respuesta de los conos: no linealidad. Fig. I.3.
luminancia tiempo Propiedades de los fotorreceptores. Respuesta de los conos: no linealidad, adaptación. J. M. Valeton y D. van Norren (1983). Fig. I.4.
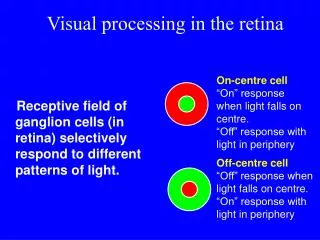
Excitación e inhibición. Respuestas on y off de una célula. Fig. I.5.
Índice de transitoriedad. Maunsell, J. H. R y Gibson, J. R (1992); [adaptado de Schiller, P. H y Malpeli J. G. (1978, 1979)]. NP1 NP2 Fig. I.6.
40 40 Magno Parvo 35 35 30 30 25 25 Número de células Número de células 20 20 15 15 10 10 5 5 0 0 0 1 2 3 4 5 6 0 1 2 3 4 5 6 Latencia de Latencia de conducción (ms) conducción (ms) Velocidad de conducción. De Schiller, P. H. y Malpeli, J. G. (1978, 1979). “desde que se produce una señal en el quiasma óptico hasta que aparece el primer pico en una célula del LGN Fig. I.7.
Propiedades temporales de M y P. Índice de transitoriedad: Magno mayores, Parvo menores Latencia: Magno cortas, Parvo más largas Velocidad de conducción (del axón): Magno altas, Parvo menores. Campo receptivo temporal (ver más adelante): Magno bifásicos, Parvo más monofásicos
P M K Sensibilidad espectral: células oponentes y no oponentes. De Valois et al. (1966) ? Fig. I.8.
x x Concepto de campo receptivo. Modelo de Rodiek. CR(r,t,l)=f(r).h(t).S(l) Fig. I.9.
Respuestas a un estímulo en diferentes regiones del campo receptivo. Fig. I.11.
Tipo I (centro L-on) Tipo II (S on) Para identificar los inputs hay que mapear el campo receptivo con los estímulos adecuados. Wiesel, T.N y Hubel, D.H. (1966). Log respuesta Fig. I.12.
+L+M -L-M M +M -L-M +L+M +L -L -M -M P -L DM=0 +L +M DL=0 DL=DM=0 S-(L+M) (L+M)-S K DS=0 Fig. I.13. Mapas de inputs y sensibilidad espacial. Tipo III Tipo I Tipo II
Nótese que para que la oponencia se manifieste es necesario que la luz llene todo el campo receptivo. Respuesta a diferentes estímulos en un campo receptivo cuyo mapa de inputs es de Tipo I (por ej. +L/-M). Fig. I.14.
Al menos en la fovea, el centro del campo receptivo recibe señal de un sólo cono. Campo receptivo de una célula oponente en la fovea. P. Lennie (2001). Fig. I.15.
KC ? l = l × 2 S (r , ) S ( ) A exp {-(r/s ) } C C C C l = l × 2 S (r , ) S ( ) A exp {-(r/s ) } P P P P KP Rodiek Modelo de Rodiek en una célula de Tipo I (+L/-M), pero no en la fovea. Fig. I.16.
Nótese que cuando el cambio es puramente cromático, elantagonismo del campo receptivo se convierte en sinergismo. Estímulos óptimos en una célula de Tipo I (+L-/M), a partir del punto neutro. DY<0 DY>0 Cambios de luminancia Dl<0 Cambios de color Dl>0 Fig. I.17.
+L+M -L-M M +M -L-M +L+M +L Tipo III -L -M -M P -L D Tipo I +L +M D S-(L+M) (L+M)-S K Tipo II CRs para cambios de lum y col (a partir del punto neutro). Fig. I.18.
+L+M -L-M M +M -L-M +L+M +L Tipo III -L -M -M P -L Tipo I +L +M S-(L+M) (L+M)-S K Tipo II Estímulos óptimos (a partir del punto neutro). Fig. I.18.
Respuesta de una célula de Tipo I (+M/-L) a una barra. De Valois, R. L. et al. (1977). +M +M Fig. I.19.
DY<0 DY<0 DY>0 3 1 3 1 2 2 CSFs acromática. Derrington, A.M. y Lennie, P. (1983). Fig. I.21a.
Magno Tipo I Las células M tienen sensibilidad de pico más alta, pero las células P tienen mejor resolución. CSFs Magno vs Parvo. Derrington, A. M. y Lennie, P. (1983). Fig. I.21b.
CSFs cromática y acromática de una célula de Tipo I. De von BlancKensee (1981). 1 1 2 2 3 3 Fig. I.22.
centro periferia Las células M tienen los CRs temporalmente bifásicos, mientras que los de las células P son más monofásicos. Pero, realmente, el espacio y el tiempo no son separables: campo receptivo espacio-temporal. Fig. I.25.
frecuencia temporal CSFs temporales cromática (izq.), acromática Parvo-Tipo I (centro ) y acromática Magno (der.). Lee, B. B. et al. (1994) Fig. I.23.