Download
1 / 53
530 likes | 880 Views
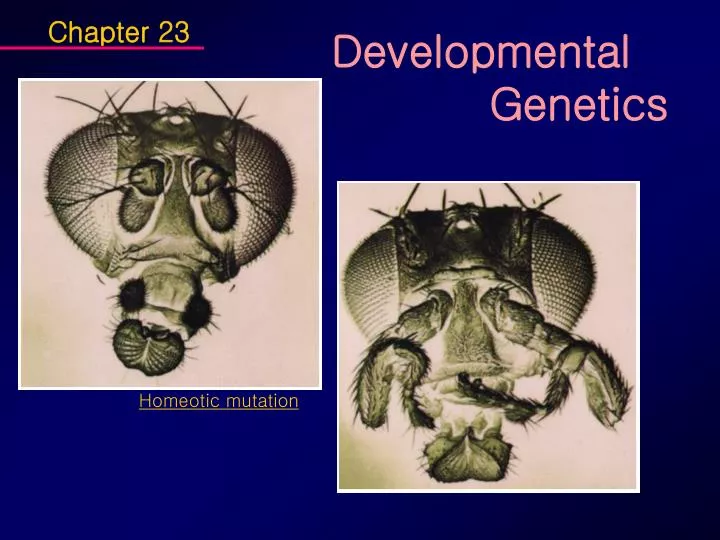
Chapter 23. Developmental Genetics. Homeotic mutation. Central themes of developmental genetics - pattern formation, construction of complex form, operation through developmental program Logic of building the body plan

E N D
Chapter 23 Developmental Genetics Homeotic mutation
Central themes of developmental genetics - pattern formation, construction of complex form, operation through developmental program Logic of building the body plan - Cell fate : differentiate into particular kinds of cells : positional information developmental field - Fate refinement : asymmetric division different regulatory instruction : decision by neighboring or paracrine signals - Specify fate option of cells of a given cell lineage in step by step manner : totipotent fate refinement cell lineage Major decisions in building the embryo - simple binary decisions : separation of the germ line from the soma : establishment of the sex of the organism - specification decisions : establishment of the positional information : anterior-posterior axis segment dorsal-ventral axis germ layer : production of the various organs, tissues, systems
Applying regulatory mechanisms to developmental decisions - Simple developmental pathway : the concentration of some key molecules determine the “on” or “off” binary choice - making a pathway decision and subsequently remembering that decision are both key to cell fate commitment Decision making in developmental pathway
- Simple developmental pathway : During development, regulatory mechanisms at any of level can control the production of active protein products Regulation of protein activity
Gene regulation at levels other than transcription Initiation:examples Tissue-specific regulation at the level of DNA structure Tissue-specific amplification of the number of copies of a gene leads to high levels of gene expression in that tissue : Eggshell gene copy number is increased by somatic DNA rearrangement only in the follicle cell : each ACE region contains an amplification control element act as a special site for the initiation of tissue-specific DNA replication in follicle cells of the ovary Structure of the two eggshell-gene clusters in Drosophila melanigaster
Transcript processing and tissue-specific regulation The production of an active protein can be regulated by controlling the pattern of splicing of an initial transcript into an mRNA Germ line specific expression of Drosophila P elements: examples of tissue-specific regulation by RNA splicing a) Somatic and germ line mRNA structure for the wild type P element b) Modified P element transgene, P {2, 3}; in all tissues, transposase is active
Transcript processing and tissue-specific regulation P transposase derived from a wild type P element: P element is inactive in soma (red eye) P transposase derives from the P {2,3} transgene: P transposase is active in soma (red and white) Somatic expression of P transposase in Drosophila containing P(w+) transposon
Posttranscriptional regulation * A certain sequences of 3’UTR of mRNA - regulate the degradation or the translation efficiency of mRNA - target sites for proteins that digest mRNA molecule or that block their translation * A 3’UTR sequences interact with a regulatory RNA molecule ex) in C. elegans - premature adult development, or reiterated, producing delayed adulthood - RNA product of the lin-4 repress translation of lin-14 mRNA (complementarity between lin-4 RNA and 3’UTR of lin-14) * A 3’UTR act as sites for anchoring an mRNA to particular structures within a cell Regulatory instructions are also contained within noncoding regions of mRNAs
Posttranslational regulation * Enzymatic modifications of proteins - phosphorylation - acetylation….. * Multiprotein complexes - protein-protein interactions
Binary fate decisions: pathways of sex determination * In many species, sex determination is associated with the inheritance of a heteromorphic chromosome pair in one sex (XX-XY system) * The XX-XY system of flies and mammals arose independently
Drosophila sex determination: every cell for itself Every cell in Drosophila independently determines its sex Phenotypic consequences of different x-chromosome-to-autosome ratios Drosophila n = 4 one sex chromosome three different autosomes X : A > 1 or = 1 -> female X : A < 0.5 or = 0.5 -> male (A : number of autosome set)
Basics of the regulatory pathway Sexual phenotype is carried out by a master regulatory switch and several downstream sex-specific genes The pathway of sex determination and differentiation in drosophila
Regulatory switch; the activity of Sxl (Sex lethal) protein 1. Setting the switch in the “on” or “off” position <The initiation & maintenance of the Sxl switch> - NUM (numerator gene in X ch.) - DEM (denominator gene in autosome) - NUM and DEM (both bHLH) forms dimers at random - Only NUM-NUM dimers forms active transcription factor
Regulatory switch 2. Maintaining the switch in a stable position - The Sxl gene has two promoters :- PE is active only early in embryogenesis :- PL is active in every cell for the remainder of the life cycle - SXL protein is an RNA-binding protein that alters the splicing of the nascent Sxl transcript coming from late promoter - Feedback or autoregulatory loop controlled at the level of RNA splicing :- maintains SXL activity throughout development in flies with an X : A ration of 1.0 Autoregulatory loop exemplifies how an early developmental decision can be “remembered” for the rest of development, even after the initial signals that established the decision have long disappeared
Regulatory switch 3. Propagating the decision Two forms of tra mRNA Different dsx mRNA are produced in both sexes Alternative splicing of tra and dsx transcripts
Mutational analysis of Drosophila sex determination “Treasure your exceptions” * The effect of null mutation - transform female into phenotypic male - Sis-b (sisterless-b); a numerator gene - Sxl (sex-lethal) and tra (transformer); the RNA splicing regulators - Must be active for female development * Dsx (doublesex) - Dsx-/- leads to the production of flies that simultaneously have male and female attributes
Sex determination in mammals: coordinated control by the endocrine system ;- the presence or absence of a Y chromosome Mammalian reproductive development and endocrine organ control The embryonic genital ridge consists of a medulla surrounded by a cortex - Female germ cells migrate into the cortex & become organized into a ovary - Male germ cells migrate into the medulla & become organized into a testis In the initial urogenital organization at the indifferent gonad stage, precursors of both male (Wolffian) and female (Mullerian) ducts are present
Mammalian reproductive development and endocrine organ control * It is the presence or absence of a testis that determines the sexual phenotype, through the endocrine release of testosterone (in XY embryos lacking the androgen receptor, development proceeds along a completely female pathway even though the embryos have testes) Setting the switch in the “on” or “off” position Testis-determining factor on Y chromosome (TDF in human, Tdy in mice), as same as SRY (human) –Sry (mice) gene The wild type XY individual has SRY gene, which activates male shunt pathway Normal XX individual lacking SRY remains in the female default pathway
Mutational analysis of mammalian sex determination Sex reversal - Sex reversed XX individuals are phenotypic males and carry a fragment of the Y chromosome in their genomes sex reversal on Y gene (SRY) A molecular map of the distal part of the short arm of the human Y chromosome
A transgenic mouse that proves Sry can cause the sex-reversal syndrome PCR of genomic DNA shows that Mouse 33.13 lacks a DNA marker (Zfy-1) for Y chromosome but, contains the Sry transgene The external genitalia of sex-reversed XX transgene mice 33.13 and normal XY male sib (33.17)
Binary fate decision: the germ line versus the soma In making the decision of germ line versus soma, the embryo exploits its machinery for creating asymmetries-the cytoskeleton, to localize a germ-line determinant to a subset of early embryonic cells
Cytoskeleton of the cell Intermediate filament (vimentin) Microtubules (tubulin) Microfilament (actin) Roles: control of the location of the mitotic cleavage plane control of the cell shape directed transport of molecules and organelles Cytoskeletal rods are polar structures Different cytoskeletal system in the same cell
Intrinsic asymmetry of cytoskeletal filaments The cytoskeleton serves as a highway system for the directed movement of subcellular particles and organelles ;- polarity is essential Polarity of subunits in an actin microfilament The distribution of tubulin in an interphase animal fibroblast
Electron micrograph of two small vesicles attached to a microtubule Kinesin attaches and moves the cargoes along the microtubules A diagram of the kinesin protein Movement of vesicles along microtubules
Localizing determinants through cytoskeletal asymmetries: the germ line The early development of C. elegans, showing the early divisions of the zygote The asymmetric distribution of P granules is microfilament dependent
In drosophila, microtubles provide the essential asymmetry of germ-line determinants Pole-cell formation at the syncitial stage of the early Drosophila embryo Eggshell removed Drosophila embryo and longitudinal section
Forming complex pattern: establishing positional information Mutational analysis of early Drosophila development Genetic assays for recessive zygotic and maternal effect mutations The phenotypes of offspring are purely a manifestation of their own genotype The phenotypes of offspring are purely a manifestation of their mother’s genotype
Cytoskeletal asymmetries and the Drosophila anterior-posterior axis Positional information along the A-P axis of Drosophila embryo; creation of concentration gradients of two transcription factors (the BCD and HB-M proteins) BCD protein (bicoid gene product) steeper gradient in the early embryos HB-M protein (hunchback gene product) shallower, longer gradient
Localization of mRNA within a cell is accomplished by anchoring the mRNAs to one end of polarized cytoskeleton chains * Bicoid mRNA is tethered to the – ends of microtubules (anterior pole) * hb-m mRNA is uniformly distributed in the embryo - nos mRNA is localized at the posterior pole - NOS protein specifically block translation of hb-m mRNA - produce the shallow A-P gradient of HB-M protein bcd mRNA BCD protein nos mRNA NOS protein The expression of the localized A-P determinant
* There are specific microtubule-association sequences located within the 3’UTRs - bcd mRNA 3’UTR localization sequences are bound a protein that can also bind the – ends of the microtubules The effect of replacement of the 3’UTR of the nanos mRNA with the 3’UTR of the bicoid mRNA on mRNA localization and embryonic phenotype
Studying the BCD gradient Genetic changes in the bcd gene alter anterior fate Concentration of BCD protein affects A-P cell fates The position of the cephalic furrow arises farther forward the posterior according to bcd+ gene dosage Exoskeletons of larvae derived from wild type and bcd maternal effect lethal mutant mothers Left : wild type, normal phenotype Right : bcd, anterior head and thoracic structures missing
Studying the BCD gradient Bcd mRNA can completely substitute for the anterior determinant activity of anterior cytoplasm The bcd anteriorless mutant phenotype can be rescued by wild type cytoplasm or purified bcd+ mRNA
Cell-cell signaling and the Drosophila dorsal-ventral axis Positional information can be established through cell-cell signaling by means of a concentration gradient of a secreted molecule The D-V positional information : DL protein activity The distribution of DL in response to the SPZ signal (spaetzle gene product) DL protein is in the nucleus ventrally, throughout the cell laterally and in the cytoplasm dorsally Dl; dorsal Cact; cactus Spz; spaetzle A mature oocyte
Cell-cell signaling and the Drosophila dorsal-ventral axis The signaling pathway that leads to the gradient of nuclear versus cytoplasmic localization of DL proteins
The two classes of positional information II. Formation of a concentration gradient of an extracellular diffusible molecule I. Localization of mRNAs within a cell Unicellular field Multicellular field Morphogen; concentration-dependent determinants of form
Forming complex pattern: utilizing positional information to establish cell fates Initial interpretation of positional information • Positional information ; a gradient of transcription factor activities • (AP axis: BCD & HB-M, DV axis: DL) • Activation of cardinal genes - the first genes to respond to the maternally supplied positional information - the zygotically expressed genes
Review of Drosophila embryology A syncitium-stage embryo, common cytoplasm toward the periphery and the central yolk-field region A cellular blastoderm embryo, columnar cells Changes during cellularization
Review of Drosophila embryology 3-hour Drosophila embryo 10-hour Drosophila embryo Newly hatched larva Segmentation identity of cells along the A-P axis is already fated early in development
* A-P cardinal genes: gap genes * The gap genes can be expressed in a series of distinct domains by having promoters that are sensitive to the concentrations of A-P transcription factors
Early blastoderm expression patterns of protein from three gap genes: hb-z, kr, and kni Early embryonic expression pattern of gap genes
Refining fate assignments through transcription-factor interactions * The gap genes activate a set of secondary A-P patterning genes ; pair-rule genes in a repeating pattern of seven stripes the correct number of segments Late blastoderm expression patterns of protein from two pair-rule genes: ftz (gray), and eve (brown) * There is a hierarchy within the pair-rule genes ; combination of pair-rule genes transcriptional regulation of the segment-polarity genes (the correct number of segments) * Repeating expression pattern of primary pair-rule genes ? Regulatory element complexity of the primary pair-rule genes turns an asymmetric (gap gene) expression pattern into a repeating one
* The establishment of segmental identity ; homeotic gene complexes - ANT-C (segmental identity in head and anterior thorax) - BX-C (segmental identity in posterior thorax and abdomen) * Homeosis : the conversion of one body part into another Wild type, one copy T2 one copy T3 A bithorax triple mutant homozygote transforms T3 into a second copy of T2 Homeotic gene transformation of the third thoracic segment (T3) of Drosophila into an extra second thoracic segment (T2)
Homeotic gene-encoded protein expression pattern in Drosophila SCR ANTP UBX ABD-B Linear arrangement of the corresponding genes along chromosome 3 Segment identity is established through asymmetric gap-gene expression patterns that deploy and asymmetric pattern of homeotic gene expression
A cascade of regulatory events Hierarchical cascade that activates the elements forming the A-P segmentation pattern in drosophila
Additional aspects of pattern formation Memory systems for remembering cell fate Two types of positive feedback loops in maintaining the level of activity of transcription factors determining cell fate The transcription factor binds to an enhancer of its own gene, maintaining its transcription Each adjacent cell sends out a signal that activates receptors, signal transduction pathways, and transcription factors (TF) expression in the other cell (cell-cell interactions) When the fate of a cell lineage has been established, it must be remembered
Ensuring that all fates are allocated: decisions by committee Cell-cell interactions: one type is the ability of one cell to induce a developmental commitment in one neighbor of many, the other is the ability of one cell to inhibit its neighbors from adopting its fate. Adult Caenorhabditis elegans
The production of the C. elegans vulva from the equivalence group by cell-cell interactions Fate allocation can be made through a combination of inductive and lateral inhibitory interactions between cells
Developmental pathways are composed of plug and play modules Differences in the developmental context of different cell lineages -that is, the transcription factors active in these cells- permit different inputs to, and outputs from, a given developmental circuit Different developmental decisions are made by using mixed and matched combinations of pathway components
The many parallels in vertebrate and insect pattern formation The similarity between the mammalian homeobox gene cluster (Hox complex) and insect ANT-C and BX-C homeotic gene cluster (HOM-C) The comparative anatomy of the HOM-C and Hox gene clusters paralogous The expression domains and regions of the Drosophila and mouse embryos : the arrangements of the homeotic genes is colinear with their spatial pattern of expression
Each Hox gene is expressed in a continuous block beginning at a Specific anterior limit and running posteriorly to the end of the developing vertebral column The RNA expression pattern of three mouse Hox genes in the vertebral column of a sectioned 12.5-day-old mouse embryo: the anterior limit of each of the expression pattern is different



















