Download

1 / 39
410 likes | 1.04k Views
Mechanism of Signal Transduction by Rhodopsin as a Model GPCR. by Basak Isin. OUTLINE. G Protein-Coupled-Receptors (GPCR) Aim of the project Gaussian Network Model (GNM) Anisotropic Network Model (ANM) Results and Discussion Conclusion and Summary Future Plans. GPCRs.

E N D
Mechanism of Signal Transduction by Rhodopsin as a Model GPCR by Basak Isin
OUTLINE • G Protein-Coupled-Receptors (GPCR) • Aim of the project • Gaussian Network Model (GNM) • Anisotropic Network Model (ANM) • Results and Discussion • Conclusion and Summary • Future Plans
GPCRs • the largest superfamily of cell surface receptors • seven helices - their signature motif • involved in a number of clinically important ligand/receptor processes. • bind ligands from the cell exterior, which induce a conformational change in the cytoplasmic face of the receptor, enabling binding of the G protein. • couple to heterotrimeric G proteins to convert an extracellular signal into an intracellular signal. • significant drug targets. 50-60% of approved drugs target members of the GPCR family.
RHODOPSINthe first 3-dimensional molecular model for a GPCR cytoplasmic region 8 4 1 6 5 2 3 7 extracellular region located in the outer segments of rod photoreceptor cells in the retina responds to environmental signals, i.e., photons initiates intracellular processes that result in an electrical signal processed by the visual system.
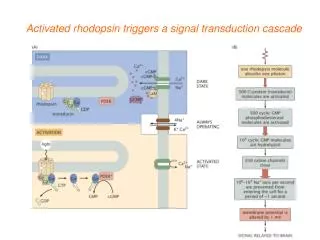
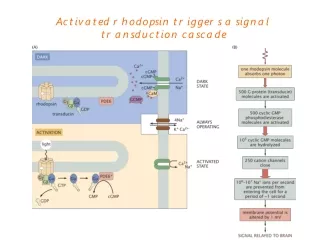
LIGHT ACTIVATION Rhodopsin Metarhodopsin II 11-cis retinal All-trans retinal
SNAKE LIKE REPRESENTATION • Glycosylation at 2 asparagines • 2 Cys(110/187) form sulfide bridge • Lys296 at H7 is covalently attached to • 11-cis retinal (protonated Schiff base) • Glu113 at H3 is the counterion to Schiff base • Palmitate attached to 2 C-terminal Cys • Ser334, 338 & 343 are major sites for • phosphorylation
Microdomains • the Asp/Glu Arg Tyr (ERY) motif at the cytoplasmic end of helix 3 • the Asn Pro X X Tyr(NPXXY) motif in helix 7 (Okada et al., 2002) • the X1BBX2X3B motif at cytoplasmic end of helix 6 (B, basic; X, non-basic) (Ballesteros et al., 1998), • an ionic interaction between the ligand and the receptor at the retinal Schiff base Lys296 and the Schiff base counterion the Glu113 (Cohen et al., 1993), • the Asn-Asp interaction between helices 1 and 2, respectively • the aromatic cluster surrounding the ligand binding pockets (Visiers et al., 2002)
AIM OF THE PROJECT Understanding the mechanism of activation of rhodopsin as a model GPCR and its interaction with Gt by structure-functions analysis using the GNM and its extension, the ANM.
Gaussian Network Model(GNM) For estimating the dynamic characteristics of biomolecular structures based on atomic coordinates in the native conformation • Elastic network • Virtual bond representation • No distinction between nonbonded and bonded neighbors • The interactions between residues in close proximity represented by harmonic potentials with a uniform spring constant
19 20 18 21 22 2 3 1 4 8 23 5 CONSTRUCTION OF KIRCHHOFF MATRIX 7A 1 2 3 4……18 19 20 21….N 3 -1 -1 -1 -1…..-1 -1 -1…-1….0
G = U L UT G-1 = U L-1UT The fluctuations associated with kth mode = (3kBT/ ) (k-1[uk]i [uk]j) U: orthogonal (NxN) matrix uk: kth eigenvector of Γ (1 ≤ k ≤ N) (shapes of corresponding mode of motion) L: diagonal matrix with eigenvalues (lk) l1 = 0 < l2 < …< lN frequency of modes
a 27-33 26-37 6-11 222-230 14-20 b d c Global Motions Slow Modes the mechanism of the motion relevant to biological function. The maxima of the slow mode curves indicate the most flexible regions of the molecule. Identifies the hinge region that are important for biological function.
RESULTS AND DISCUSSION Comparison of theoretically calculated all modes with B- factors found by X-ray crystallography Science, Palcwezski et. al, 2000
1 2 7 5 6 3 H2 H5 H6 H7 H3 H4 H1 First Mode of GNM The color codes are green, cyan, blue, magenta, pink and yellow in the order of increasing mobility.
a R esidue number Residue number CROSS CORRELATIONOf GNM Residue number Residue number Residue number
Fast Modes Critically important ones for the overall stability of the molecule and evolutionarily conserved High Frequency Modes Asn55
a Front View Back View b 6 8 6 8 8 2 7 2 7 5 2 1 4 3 1 3 5 7 2 1 6 1 6 5 4 4 5 3 7 ANM results ANM results Wild type ANM • the three components of the inter-residue separation vectors obey Gaussian dynamics. • involves the inversion of a 3N x 3N Hessian matrix H that replaces the N x N Kirchhoff matrix (Doruker et al., 2000, Atilgan et al., 2001).
MOVIES FRONTBACK
b 4 4 1 2 2 3 3 1 8 5 7 5 6 6 8 7 wild type ANMb ANMa 8 8 8 4 3 3 4 4 5 5 1 7 7 5 6 1 7 6 6 2 1 2 2 3 TWO CONFORMATIONS ANMa MM(199): E=-5531.2 ANMb MM(273): E=-5235.41
Top-248 Cross-sections from the Top 248-255
255-263 264-273
273-277 bottom
Rij H1 H2 H3 H4 H5 H6 H7 H8
ALL ATOM MODEL • to see the side chain motions • to study the microdomains • Adding the fluctutations of C to every atom • in the PDB structure • Energy minimize the structure
E-MIN TRANS-RETINAL NO RETINAL CIS-RETINAL
MICRODOMAIN1-ERY ANM WILD TYPE
CONCLUSION • The rhodopsin ground state structure is predisposed for the functional conformational changes leading to the opening of the helical bundle, thereby revealing the mechanism for this process. • The mechanism for the observed opening of the helical bundle is mediated by a torsional rotation of the molecule centered on Helix 3. • The cytoplasmic end of helix 4 moves away from the cytoplasmic end of helix 3, the most flexible region in helix 3, and stretches the cytoplasmic loop 2. • Helix 6 is rotates while simultaneously elongating, comparable to a turning screw. • Furthermore, they suggest a mechanism for the activation of the G-protein. • The screwing motion of helix 6 may provide a mechanical trigger for conformational changes in the G-protein which lead to GDP/GTP exchange.
SPECIFIC AIM 1A.The mechanism of activation of Rhodopsin by improved GNM/ANM Analysis 1. Side Chain Interactions 2. Water Molecules 3. Chromophore 4. Missing loops 5. Lipid molecules B. Chromophore binding pocketC. Surface exposure after opening of the helical bundle during activationD. Rhodopsin oligomerization(Fotiadis et al, 2003 Nature)E. Analysis of multiple modes of the GNM and ANM
SPECIFIC AIM 2Interaction of rhodopsin with Gt • interaction surface for rhodopsin- Gt complex by electrostatic surface maps of the active form of rhodopsin found by ANM analysis and Gt. • Analysis of GDP Bound form of Heterotrimeric Gt by GNM and ANM.
SPECIFIC AIM 3 Extension of the mechanism of Activation to other GPCRs • by exploring the importance of hinge regions by sequence alignment. • by applying the GNM and ANM analysis to other members of the family whose structures are determined theoretically. Metabotropic Glutamate receptors Adrenergic receptors
Thanks to Dr. Judith Klein -Seetharaman Dr. A.J. Rader Dr Mike Cascio Dr Billy Day Dr Christine Milcarek Dr Hagai Meirovitch Dr Tom Smithgall Dr. Ivet Bahar Post-Docs: Dr. Rajan Munshi Dr. Dror Tobi Graduate Students Elife Zerrin Bagci Shann-Ching Chen Chris Myers Alpay Temiz Lee-Wei Yang System Administrators Dr. Rob Bell Mark Holliman Administrators Joseph Bahar Nancy Gehenio
Summary and Conclusion The elastic network models (GNM and ANM) are tools to explore the dynamics of proteins and to determine the criticallyimportant sites.These are classified in two categories: • The first category comprises the residues that are important for coordinating the cooperative motions of the overall molecule. These are identified from the minima of the global mode shapes. Their mutation can impede function. • The second one consists of residues experiencing an extremely strong coupling to their close neighbors, and thereby undergoing the highest frequency/smallest amplitude vibrations. Their mutation can impede stability. • Both groups of residues are expected to be evolutionarily conserved, the former for function requirements, and the latter for folding and stability
Experiments to validate the calculations • Site directed spin labeling combined with EPR • Cysteine scaning mutagenesis: -Reactivity and solvent accesibility of the sulfhydryl groups to 4-PDS. Absorbance of 4-TP at 323nm -Disulfide exchange of thiopyridinyl-derivatives of rhodopsin by sulfhydryl reagents (R-SH) both in dark and after illumination. -Disulfide bond formation in double cysteine mutation assay. Rate of cysteine bond formation is a measure of proximity of the mutant. In addition, sulfur bridges can inhibit light activation. This can show the necessary movements of helices to form MetaII. • Site directed19F labeling for NMR study: single cysteine mutants followed by the attachment of TET (CF3-CH2-S) attachment. Shifts in dark and light NMR spectra of the mutants shows the movements of residues. • Antibody binding experiments: Antibodies which bind to Meta II but not rhodopsin in the dark.
Experiments to validate the calculationsRhodopsin-Gt Interaction • Gt activation by fluoresence spectroscopy: After GTPS addition, increase in fluoresence results from exposure of Trp207 in Gt. when Gt is activated. • Assay of Meta II-Gt complex: Flash photolysis (light scattering). A flash induced light scattering increases over time in the presence of binding but not in the absence of Gt. This signal reflects the binding of Gt to R. In the presence of Gt and GTP, a flash produces a decrease of scattering intensity due to a loss of scattering mass. • Nucleotide release assay for GDP release ability. Samples are filtered through a nitrocellulose membrane. The amount of [32P]GDP is filtered and quantitated. • Peptide competition assays.