Download
1 / 1
10 likes | 39 Views
Study on genotypic similarity among Metarhizium anisopliae isolates using AFLP technique. Results indicate variations among isolates. Research aims to understand genetic diversity and relationships between different M. anisopliae varieties.

E N D
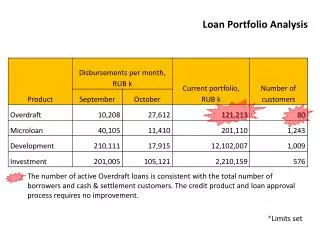
PDAY media ARSEF 324 ARSEF 2575 ARSEF 3341 ARSEF 3609 ARSEF 5749 ATCC 10074 0.0060% 0.0005% 0.0020% 0.0010% 0.0001% 0.0004% 0.0003% 0.0080% 0.0002% 0.0040% Control 0.0250% 0.0300% 0.0100% 0.0150% 0.0200% 14 day old cultures AFLP Analysis of Metarhizium AnisopliaeIsolates E. K. K. Fernandes, M. P. Miller, J. Orwin, D. E. N. Rangel, D. W. Roberts Department of Biology, Utah State University, Logan, UT 84322-5305, USA. Departamento de Parasitologia Animal, Universidade Federal Rural do Rio de Janeiro, RJ, Brasil distilled water up to 50l. After digestion, we added 10 l of a solution containing: 0.5 l EcoRI-adapters (10 mM) and 0.5 l MseI-adapters (100 mM), 1 unit T4 DNA-ligase, 1.2 mM ATP, 1 l 10 RL buffer, and double distilled water up to 10 l. The incubation was continued for 3 h at 37 ºC. After ligation, the reaction mixture was stored at –20 ºC. A PCR pre-amplification was carried out containing 6 l ligated DNA, 0.5 l EcoA (10 mM), 0.5 l MseA (10 mM), 2 l dNTP (5 mM), 3 l MgCl2 (25 mM), 1 unit Taq polymerase, 5 l 10 Taq polymerase buffer, and sterile double distilled water up to 50 l. The PCR pre-amplification cycle included a 2-min 72 ºC step, 29 cycles of 94 ºC for 30 sec, 98 ºC for 30 sec and 72 ºC for 1 min. Ten l of pre-amplification products and 2 l of a light-colored loading dye were electrophoresed at 120 V on 1% agarose gel, using Tris-acetate EDTA buffer. A 100 bp DNA ladder was run on the same gel as size marker. After electrophoresis, the gels were stained with ethidium bromide and visualized under ultraviolet light. Multilocus AFLP profiles were obtained from three additional PCR amplifications. Primers used were: A for MseI and AGC for EcoRI primer extension; A for MseI and AGG for EcoRI primer extension; and A for MseI and ACC for EcoRI primer extension. For each primer combination, a PCR amplification was carried out containing 2.5 l PCR pre-amplification product diluted 1:10 in water, 0.05 l Eco primer (10 mM), 0.2 l Mse primer (10 mM), 0.4 l dNTP (5 mM),0.6 l MgCl2 (25 mM), 0.5 unit Taq polymerase, 1 l 10 Taq polymerase buffer, and sterile double distilled water up to 10 l. The PCR amplification was run under the following thermalcycler parameters: 94 ºC for 2 min, then start a ‘touchdown’ PCR procedure using a 30 sec denature step at 94 ºC, a 30 sec annealing step at 65 ºC, and a 1 min extension step at 72 ºC. This procedure was repeated 9 times, each time reducing the annealing temperature by 1 ºC. The annealing temperature of the final ‘touchdown’ step was 57 ºC, followed by 30 cycles of the following conventional thermalcycler program: 94ºC for 30 sec, 56 ºC for 30 sec, 72 ºC for 30 sec. An appropriate quantity of ABIs ROX 400 size standard was added and loaded in the ABI 3100. AFLP markers between 50-400 bp were analyzed with Genographer software (freely-distributed at http://hordeum.oscs.montana.edu/genographer/). Cluster analysis was performed using NTSYS-pc vers2.1 (Exeter Software, Setauket, New York) and a combined dendrogram from all three primer pairs was derived from the similarity matrix calculated using simple match coefficient. Cluster analysis was performed using the unweighted pair-group method with arithmetic mean (UPGMA). Analyses were performed on all 11 Metarhizium anisopliae isolates. acridum are clustered in the first group andthe isolates of M. anisopliae var. anisopliae in a different group (Figure 1). Introduction Another study (not published) showed that ARSEF 3609 and isolates of M. anisopliae var. acridum (ARSEF 324 and 3341) had similar tolerance to a fungicide Dodine (Figure 5). Metarhizium anisopliae is an important entomopathogenic fungus that occurs worldwide, and some isolates are used for biological control of arthropods population. In a recent study on characterization of 100 Metarhizium cultures the isolate ARSEF 3609 was identified as M. anisopliae var. anisopliae (Driver et al. 2000). This isolate, however, has physiologic features (e.g. host hange, culture aspect, UV-B tolerance) more compatible with those of M. anisopliae var. acridum (Braga et al. 2001, Rangel et al. 2005, Alston et al. 2005). This study was conducted to determine if there is genotypic similarity among ARSEF 3609 and other isolates of M. anisopliae varieties anisopliae and acridum. Figure 3. Mean relative percentage culturability of Metarhizium strains after exposure for 4 h to irradiances of 920 and 1200 mW m-2 (Braga et al. 2001). Figure 1. AFLP dendrogram generated by Simple Matching Coefficient for all Metarhizium anisopliae isolates used in this study. The isolates F985, Green Guard and ARSEF 324, supposely are the same isolate, and have different names due to different culture histories. The AFLP results suggest that they are indeed the same isolate. Material and Methods Metarhizium anisopliae isolates The different hosts and geographic origins of the eleven isolates of Metarhizium anisopliae used in this study are listed in Table 1. The isolate ARSEF 3609 clustered with other isolates of variety acridum, even though but this isolate was considered as variety anisopliae by Driver et al. (2000) when the mitochondrial ITS1 and ITS2 region was analyzed. Nevertheless, some physiological evidences corroborate that ARSEF 3609 is an M. anisopliae var. acridum. Rangel et al. (2005) exposed conidia of 17 isolates of M. anisopliae to heat. After 8 and 12 h exposure at 45 ºC, only the isolates ARSEF 324 and 3609 had viability greater than 40% (Figure 2). The isolate ARSEF 324 (a known M. anisopliae var. acridum) andARSEF 3609 demonstrated similar thermotolerance. Table 1. Metarhizium anisopliae isolates. Their host or substrate, origin and latitude. Figure 4. Comparison of Abbott-corrected and mycosis-confirmed (sporulation observed) mortality of C. nenuphar larvae (southern population) infected with different isolates of M. anisopliae 8 days after treatment (Alston et al. 2005). Fungal cultures The isolates were cultured in flasks containing 100 ml of Potato Dextrose Broth (DIFCO). Broth cultures were shaken at 130 rpm at 25 ºC for 4 d. The fungal cells (mycelium) were collected by vacuum filtration and stored at –20 ºC. DNA extraction The fungal cells were ground in liquid nitrogen with a mortar and pestle, suspended in 1 ml sterile TE buffer and stored at –20 ºCuntil use. After defrosting, ca. 70 mg of fungal cells of each isolate was placed into 2 ml tubes and the genomic DNA was extracted using the DNeasy® Plant Mini Kit (Qiagen), following the manufacturer’s protocol. The DNA extracts were electrophoresed on 0.7% agarose gels at 120V in Tris-Acetate-EDTA buffer to estimate DNA quantity and quality. A 100 bp DNA ladder was used as a size marker.The gels were stained with ethidium bromide and visualized under ultraviolet light. AFLP DNAs were double digested for 1 h at 37 ºC with 5 units EcoRI and 5 units MseI, 5 l 10 RL buffer, and sterile double Figure 5. ARSEF (Metarhizium anisopliae) and ATCC 10074 (Aspergillus) isolates cultured on PDAY with different concentration of Dodine for 14 d. Control plates do not contain Dodine (Dettemmeir et al. in preparation). Figure 2. Mean relative percentage of germination of M. anisopliae isolates after exposure to temperatures of 40 °C (open bars) and 45 °C (closed bars) for 2, 4, 8, and 12 h (Rangel et al. 2005). References The effects of exposure to UV-B irradiances of 920 and 1200 mW m-2 on conidial relative percentage culturability of the 30 Metarhizium isolates was evaluated by Braga et al. (2001). After 4 h exposure, ARSEF 324 and ARSEF 3609 exhibited the highest relative culturability at both irradiance levels (Figure 3). Alston et al. (2005) observed that ARSEF 3609 was not pathogenic to plum curculio (Figure 4). M. anisopliae var. acridum, at the recommended dose for control of Orthoptera, under field conditions, was not pathogenic to other insect groups (Peveling and Demba, 1997; Goettel and Jaronski, 1997 and Milner et al. 2002). 1- Alston, D. G.; Rangel, D. E. N.; Lacey, L. A.; Golez, H. G.; Kim, J. J., Roberts, D. W. 2005. Evaluation of novel fungal and nematode isolates for control of Conotrachelus nenuphar (Coleoptera: Curculionidae) larvae. Biological Control: 35, 163-171. 2- Braga, G. U. L.; Flint, S. D.; Miller, C. D.; Anderson, A. J.; Roberts, D. W. 2001. Variability in response to UV-B among species and strains of Metarhizium isolated from sites at latitudes from 61 ºN to 54 ºS. Journal of Invertebrate Pathology: 78, 98-108. 3- Driver, F.; Milner, R. J.; Trueman, J. W. H. 2000. A taxonomic revision of Metarhizium based on a phylogenetic analysis of rDNA sequence data. Mycological Research: 104, 134-150. 4- Goettel, M. S. and Jaronski, S. T. 1997. Safety and registration of microbial agents for control of grasshoppers and locusts. Memoirs of the Entomological Society of Canada: 171, 83-99. 5- Milner, R. J., Baker, G. L., Cliff, A. D. 2002. Effect of parasitism by Tricopsideaoestracea Westwood (Diptera: Nemestrinidae) on the susceptibility of Phaulacridium vittatum (Sjostedt) (Orthoptera: Acrididae) to infection by Metarhizium anisopliae var. acridum. J. Entomol.: 41, 82-87. 6- Peveling, R., Demba, S. A. 1997. Virulence of the entomopathogenic fungi Metarhizium flavoviride gams and rozsypal and toxicity of diflubenzuron, fenitrothion-esfenvalerate and profenofos-cypermethrin to non-target arthropods in Mauritania. Arch. Environ. Contam. Toxicol.: 32, 69-79. 4- Rangel D. E. N.; Braga, G. U. L.; Anderson, A. J.; Roberts, D. W. 2005. Variability in conidial thermotolerance of Metarhizium anisopliae isolates from different geographic origins. Journal of Invertebrate Pathology: 88, 116-125. Results and Discussions There were 123 polymorphic bands produced by all three primers pairs. A combined dendrogram for all three banding patterns revealed two clusters of isolates. The first group with 90% of similarity and the other one with 65% of similarity. Interesting is the fact that the isolates of M. anisopliae var. Acknowledgments This research was supported by grants from the Utah Department of Agriculture and Food, the Community/University Research Initiative of Utah State University and PhD scholarships from National Council for Scientific and technological Development (CNPq) of Brazil for Everton K. K. Fernandes and Drauzio E. N. Rangel.