Download
1 / 1
10 likes | 137 Views
ADAPTATION AND RE-EDUCATION OF THE MOTOR SYSTEM : A BEHAVIOURAL, MOLECULAR AND CELLLULAR STUDY. Soude J. (1), Grondard C. (1), Launay T. (1), Gasc J. P. (2), Vidal P. P. (1), Gallien C.L. (1), Chanoine C. (1) & Charbonnier F. (1)

E N D
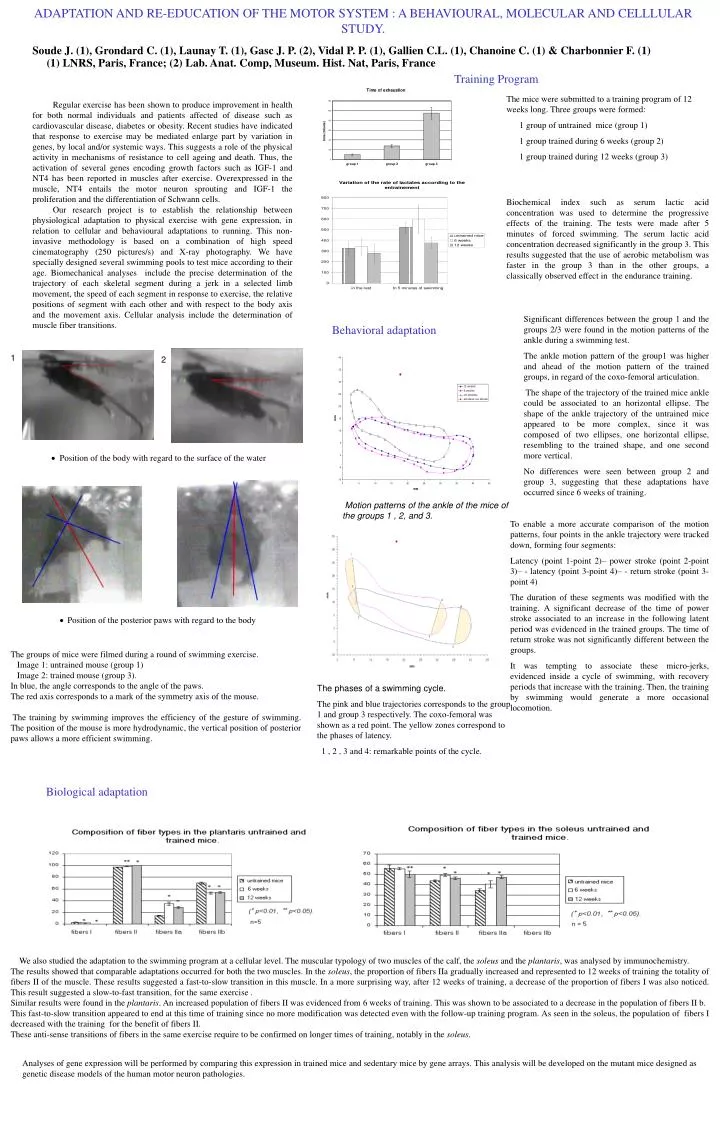
ADAPTATION AND RE-EDUCATION OF THE MOTOR SYSTEM : A BEHAVIOURAL, MOLECULAR AND CELLLULAR STUDY. Soude J. (1), Grondard C. (1), Launay T. (1), Gasc J. P. (2), Vidal P. P. (1), Gallien C.L. (1), Chanoine C. (1) & Charbonnier F. (1) (1) LNRS, Paris, France; (2) Lab. Anat. Comp, Museum. Hist. Nat, Paris, France Training Program The mice were submitted to a training program of 12 weeks long. Three groups were formed: 1 group of untrained mice (group 1) 1 group trained during 6 weeks (group 2) 1 group trained during 12 weeks (group 3) Regular exercise has been shown to produce improvement in health for both normal individuals and patients affected of disease such as cardiovascular disease, diabetes or obesity. Recent studies have indicated that response to exercise may be mediated enlarge part by variation in genes, by local and/or systemic ways. This suggests a role of the physical activity in mechanisms of resistance to cell ageing and death. Thus, the activation of several genes encoding growth factors such as IGF-1 and NT4 has been reported in muscles after exercise. Overexpressed in the muscle, NT4 entails the motor neuron sprouting and IGF-1 the proliferation and the differentiation of Schwann cells. Our research project is to establish the relationship between physiological adaptation to physical exercise with gene expression, in relation to cellular and behavioural adaptations to running. This non-invasive methodology is based on a combination of high speed cinematography (250 pictures/s) and X-ray photography. We have specially designed several swimming pools to test mice according to their age. Biomechanical analyses include the precise determination of the trajectory of each skeletal segment during a jerk in a selected limb movement, the speed of each segment in response to exercise, the relative positions of segment with each other and with respect to the body axis and the movement axis. Cellular analysis include the determination of muscle fiber transitions. Biochemical index such as serum lactic acid concentration was used to determine the progressive effects of the training. The tests were made after 5 minutes of forced swimming. The serum lactic acid concentration decreased significantly in the group 3. This results suggested that the use of aerobic metabolism was faster in the group 3 than in the other groups, a classically observed effect in the endurance training. Behavioral adaptation Significant differences between the group 1 and the groups 2/3 were found in the motion patterns of the ankle during a swimming test. The ankle motion pattern of the group1 was higher and ahead of the motion pattern of the trained groups, in regard of the coxo-femoral articulation. The shape of the trajectory of the trained mice ankle could be associated to an horizontal ellipse. The shape of the ankle trajectory of the untrained mice appeared to be more complex, since it was composed of two ellipses, one horizontal ellipse, resembling to the trained shape, and one second more vertical. No differences were seen between group 2 and group 3, suggesting that these adaptations have occurred since 6 weeks of training. 1 2 Position of the body with regard to the surface of the water Motion patterns of the ankle of the mice of the groups 1 , 2, and 3. To enable a more accurate comparison of the motion patterns, four points in the ankle trajectory were tracked down, forming four segments: Latency (point 1-point 2)– power stroke (point 2-point 3)– - latency (point 3-point 4)– - return stroke (point 3-point 4) The duration of these segments was modified with the training. A significant decrease of the time of power stroke associated to an increase in the following latent period was evidenced in the trained groups. The time of return stroke was not significantly different between the groups. It was tempting to associate these micro-jerks, evidenced inside a cycle of swimming, with recovery periods that increase with the training. Then, the training by swimming would generate a more occasional locomotion. Position of the posterior paws with regard to the body The groups of mice were filmed during a round of swimming exercise. Image 1: untrained mouse (group 1) Image 2: trained mouse (group 3). In blue, the angle corresponds to the angle of the paws. The red axis corresponds to a mark of the symmetry axis of the mouse. The training by swimming improves the efficiency of the gesture of swimming. The position of the mouse is more hydrodynamic, the vertical position of posterior paws allows a more efficient swimming. The phases of a swimming cycle. The pink and blue trajectories corresponds to the group 1 and group 3 respectively. The coxo-femoral was shown as a red point. The yellow zones correspond to the phases of latency. 1 , 2 , 3 and 4: remarkable points of the cycle. Biological adaptation We also studied the adaptation to the swimming program at a cellular level. The muscular typology of two muscles of the calf, the soleus and the plantaris, was analysed by immunochemistry. The results showed that comparable adaptations occurred for both the two muscles. In the soleus, the proportion of fibers IIa gradually increased and represented to 12 weeks of training the totality of fibers II of the muscle. These results suggested a fast-to-slow transition in this muscle. In a more surprising way, after 12 weeks of training, a decrease of the proportion of fibers I was also noticed. This result suggested a slow-to-fast transition, for the same exercise . Similar results were found in the plantaris. An increased population of fibers II was evidenced from 6 weeks of training. This was shown to be associated to a decrease in the population of fibers II b. This fast-to-slow transition appeared to end at this time of training since no more modification was detected even with the follow-up training program. As seen in the soleus, the population of fibers I decreased with the training for the benefit of fibers II. These anti-sense transitions of fibers in the same exercise require to be confirmed on longer times of training, notably in the soleus. Analyses of gene expression will be performed by comparing this expression in trained mice and sedentary mice by gene arrays. This analysis will be developed on the mutant mice designed as genetic disease models of the human motor neuron pathologies.