Download

1 / 34
340 likes | 548 Views
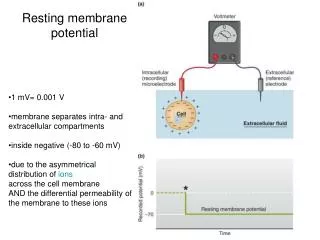
The Resting Potential. Cells are electrical batteries. Virtually all cells have a steady transmembrane voltage, the “resting potential”, across their plasma membranes. The negative pole of the battery is the interior of the cell; the positive the exterior.

E N D
Cells are electrical batteries • Virtually all cells have a steady transmembrane voltage, the “resting potential”, across their plasma membranes. • The negative pole of the battery is the interior of the cell; the positive the exterior. • All voltage values are measured relative to some baseline – in this case, we usually take the solution surrounding the cell as the ground or baseline, and so resting potential values are expressed as negative numbers. • We can measure the resting potential by inserting a metal or glass electrode across the plasma membrane, placing a second (ground) electrode near the cell surface, and connecting a voltmeter to the electrodes.
What are the sources of this electrical potential energy? • Direct contributions from pumps that move charge – this would include both the Na+/K+ pump (almost all cells) and the V-type H+ ATPase (restricted to a few cell types). • Diffusion potentials arising from ionic gradients
Diffusion potentials and the concept of electrochemical equilibrium • Imagine two solutions of differing ionic composition, separated by a barrier. For example, let’s let the solute be KCl and the gradient be 10:1. • Depending on the permeability properties of the barrier, there are 4 possible outcomes (but two of them are boring): KCl KCl
The non-boring outcomes • 1. barrier permeable to K+ but not to Cl-: K+ will attempt to diffuse from left to right – but very soon the pull of the left-behind Cl- will become equal to the “push” of the concentration gradient, and the system will come into electrochemical equilibrium with a net negative charge on the left side of the barrier and a net positive charge on the right side. • 2. barrier permeable to Cl- but not to K+: exactly the opposite will happen, resulting in a net negative charge on the right side and an opposing positive one on the left side. • These are equilibria, so they will persist without any energy expenditure as long as the system is not disturbed.
The boring outcomes • 1. barrier permeable to both ions: a temporary diffusion potential will exist because the diffusion coefficient of K+ and Cl- differ, but ultimately concentrations will be equal on both sides and there will be no voltage at equilibrium – boring! • 2. barrier permeable to neither ion: no change at all – very boring!
The Nernst Equation relates chemical and electrical driving forces R and T have their usual meanings, Z is the ionic charge (+1 for K+), and F is Faraday’s Number, a fudge factor that converts from coulombs (a measure of static charge) to molar units. For ease of calculation, it helps to know that if we fill in constants and convert to 10-base logs, the equation yields 55 mV of potential for every additional decade of ionic gradient at room temperature or about 60 mV at mammalian body temperature.
+60 ENa This diagram shows the Nernstian equilibrium potential values for Na+ and K+ when the concentration ratios across the membrane barrier are 1/10 for Na+ and 1/30 for K+ - these are typical values for real cells 0 mV -90EK
Possible misconceptions: typical illustrations grossly under-represent the numbers of ions, so that it seems that the cell below has more than twice as many negatively charged ions inside it as positively charged ions…
The real situation: • The charge on the membrane is generated by an extremely small charge imbalance and represents very few ions. The oppositely-charged ions clustered on the inside and outside of the membrane are such a small portion of the total number of each category of ion, that for a large neuron, if one K+ diffuses out of the cell for every 10 million K+ inside the cell, the effect is to produce a membrane potential of 100mV inside-negative!
+60 ENa Where is the resting potential in this? 0 mV -70 -90 EK
Implications of the previous slide • The resting potential cannot be explained as a pure K+ or pure Na+ diffusion potential • Neither K+ nor Na+ is in electrochemical equilibrium – K+ is close, but Na+ is way off.
The Na+/K+ pump explains the non-equilibrium distributions of Na+ and K+ • If an ion’s concentration gradient is not in agreement with what the Nernst Equation predicts, work is being done to keep the system out of equilibrium. • Na+ and K+ distributions across the plasma membrane are kept away from diffusional equilibrium by the Na+/K+ pump. The energy is provided by hydrolysis of ATP.
The magnitude and polarity of the resting potential are determined by two factors: • 1. The magnitude of the concentration gradients for Na+ and K+ between cytoplasm and extracellular fluid. • 2. The relative permeabilities of the plasma membrane to Na+ and K+.
Since the Na+ and K+ concentration gradients are opposite, you could think of the membrane potential as the outcome of a tug-of-war between the two gradients. The winner (defined as the ion that can bring the membrane potential the closest to its own equilibrium potential) is determined by the relative magnitudes of the K+ and Na+ gradients and the relative permeability of the membrane to the two ions.
K+ is the winner on both counts: its gradient is about 30/1 as compared to Na+’s 10/1, and the membranes of most cells are 50-75 times more permeable to K+ than Na+.
Leak Channels Despite the overall high resistance of the membrane, some leak channels are open in the ”resting” membrane. A few of the leak channels allow Cl- through, a few allow Na+ through, but most of the leak channels allow K+ to pass through. Given that there are leak channels, which way will each ion move through the leak channels, on average?
We can quantify the effects of the Na+ and K+ gradients • We just have to know the relative magnitudes of the concentration gradients and the relative permeabilities
The Goldman Equation describes the membrane potential in terms of gradients and permeabilities In words, the Goldman equation says: “The membrane potential is determined by the relative magnitudes of the concentration gradients, each weighted by its relative permeability.”
What ions have to appear in the Goldman equation? • To be accurate, the Goldman equation must include a term for each ion that is: • a. not at equilibrium, and • b. for which there is significant permeability • So, for those cells which actively transport Cl-, a Cl- term must be added. To do so, [Cl-]in and [Cl-]out have to be inverted relative to the cation terms, because of the charge difference.
This diagram shows the sizes of the driving forces that act on Na+ and K+ when the concentration ratios across the membrane are 1/10 for Na+ and 1/30 for K+ and the resting potential is -70 mV. +60 ENa 0 Driving force on Na+ = 130 mV mV Resting potential -70 Driving force on K+ = 20 mV -90 EK
Na+ is not conflicted! • Both the concentration gradient and the internally-negative membrane potential favor entry into the cell….
The way things look to K+ • The forces on K+ are outward, down its concentration gradient, and inward, responding to the attraction of the negative interior…
Chloride is often passively distributed….. • Cl- is driven out, repulsed by the negative charge inside, but it is driven in by its concentration gradient. The result can be that Cl- is “contented” at the resting membrane potential, with its two forces balanced.
A little review of electrical terms Ohm’s Law ( I = V/R ) is the relationship between electrical force and flow. The driving force (V or E; units of volts); this is potential energy. Resistance is R (units: ohms); conductance (G) is the inverse of resistance (units: mohs or siemens) Current (I) is in units of amps: One amp is the current that flows when the driving force is 1 volt and the resistance is one ohm (or the conductance is 1 Siemen).
Relevant membrane properties: resistance and capacitance • The lipid bilayer has a high electrical resistance (i.e., charged particles do not move easily across it) and it separates two very conductive (“salty”) solutions. • The lipid bilayer is thin (about 50 Angstroms). The thinness of the membrane allows it to store a relatively large amount of charge, i.e., have a high capacitance: very small differences in the electrical balance of charges inside the cell easily attract opposite charges to the outside of the cell.
Net current flow across the membrane is zero at rest.(or at any time when the potential is stable) An important corollary of Ohm’s law is that when the membrane potential is stable, net current flow across it is zero. If net current flow is not zero, Vmembrane has to be changing.
Current Flow causes the membrane potential to change In physiology (unlike physics), current is defined as the flow of positive charge. A net inward current is thus equivalent to flow of cation into the cell (or anion out of the cell), either of which would cause depolarization - change toward a less inside-negative membrane potential. The opposite change is hyperpolarization.
Membrane response to injected current After the injected current is turned off, the membrane potential moves pretty quickly back to the resting level: What is going on?.
Some factors that cause depolarization [K+] extracellular [Na+] extracellular Na+ permeability K+ permeability