Download

1 / 65
670 likes | 1.09k Views
RNAi applications. Michela A. Denti 30 Novembre 2006. RNA interference. Biological research Define gene functions (gene knockout) C. elegans genome RNAi projects Define biochemical pathways microarray screenings of RNAi knockouts Therapeutic treatments Cancer Viral infections

E N D
RNAi applications Michela A. Denti 30 Novembre 2006
Biological research Define gene functions (gene knockout) C. elegans genome RNAi projects Define biochemical pathways microarray screenings of RNAi knockouts Therapeutic treatments Cancer Viral infections Parasitic infections Neurodegenerative diseases Dominant genetic diseases Applications of RNAi
Studi di genetica funzionale RNA interference nei Nematodi • Modalita’ di somministrazione: • “Feeding” • “Soaking” • Iniezione
Mutante engrailed Mutante twist RNAi engrailed RNAi twist Misquitta e Paterson (1999) RNA interference in Drosophila Microiniezione di dsRNA in embrioni ..oppure Espressione in vivo di sequenze ripetute invertite omologhe al gene da silenziare
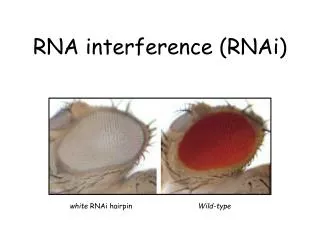
RNAi: uno strumento per inibire l’espressione genica in vivo Piante C. elegans Cellule HeLa Drosophila
RNAi: uno strumento per inibire l’espressione genica in vivo • C. elegans (Fire et al., 1998) • Drosophila (Carthew et al., 1998) • Planaria (Newmark et al., 1998) • Tripanosoma (Ullu et al., 1998) • Idra (Lohmann et al., 1999) • Zebrafish (Wargelius et al., 1999) • Topo (Wianny & Zernicka-Goetz, 2000) • “cosuppression” in piante • “quelling” in Neurospora
dsRNA induces a non-specific response in mammalian cells Cell death Kumar M, Carmichael GG. “Antisense RNA: function and fate of duplex RNA in cells of higher eukaryotes.” Microbiol Mol Biol Rev (1998) 62:1415-34
Short RNAs (<30) avoid PKR pathway Dicer siRNAs are not recognised by PKR!
19 nt duplex 2 nt 3’ overhangs Use of siRNAs in mammalian cells Transfection of siRNAs Mismatches with the target are not tolerated. Elbashir et al., (2001) Caplen et al., (2001)
Sinthetic siRNAs easy many target sequences can be tested but expensive short-term effect (depends on the cell type) in vitro synthesized and cleaved dsRNA (Dicer, RNase III) inexpensive but need to purify short-term effect DNA vectors more laborious inexpensive long-term effect 19 nt duplex 2 nt 3’ overhangs How to provide siRNAs to a cell?
Synthetic siRNAs injected in vivo McCaffrey et al., (2002) RNA interference in adult mice. Nature 418, 38-39 Injection of siRNAs in the caudal vein
Synthetic siRNAs injected in vivo Lewis et al., (2002) Efficient delivery of siRNA for inhibition of gene expression in postnatal mice. Nature Genetics 32, 107-108 EGFP transgenic mice Liver
Synthetic siRNAs injected in vivo Song et al., (2003) RNA interference targeting Fas protects mice from fulminant hepatitis. Nature Medicine 9, 347-351 • Fas-mediated apoptosis is important in hepatic damage. • Those siRNAs which knockdown Fas in hepatocytes also protect mice from fulminant hepatitis induced by an anti-Fas antibody which activates Fas (agonist)
RNA chain RNA chain base base OCH2 OCH2 O O O OH O O RNA chain RNA chain Chemical modifications • siRNAs have short intravenous life because they are degraded by nucleases and filtered by kidneys • Chemical modifications can improve their stability • Do chemical modifications impair their function? 2’-OMe CH3
RNAi in embryonic tissue Method of topical injection of esiRNAs directed electroporation and whole embryo culture Calegari F, Haubensak W, Yang D, Huttner WB and Buchholz F (2002) “Tissue-specific RNA interference in postimplantation mouse embryos with endoribonuclease-prepared short interfering RNA” Proc. Natl Acad. Sci. USA 99, 14236-40
RNAi in embryonic tissue Calegari F, Haubensak W, Yang D, Huttner WB and Buchholz F (2002) “Tissue-specific RNA interference in postimplantation mouse embryos with endoribonuclease-prepared short interfering RNA” Proc. Natl Acad. Sci. USA 99, 14236-40 Telencephalon diencephalon Knock-in mouse line expressing GFP from the Tis21 locus Expressed in neuroepithelial cells during the switch from proliferation to neurogenesis
Pol III prom TTTTT TTTTT Pol III prom Transcription sense antisense Base-pairing siRNA Vectors for siRNA expression Let the cell itself produce the siRNAs! 1)Each strand is separately transcribed Miyagishi and Taira (2002)
TTTTT Transcription shRNA Processing siRNA Vectors for siRNA expression Let the cell itself produce the siRNAs! 2) Transcription of pre-miRNA-like hairpin precursors loop Pol III prom Castanotto et al. (2002) Paul et al. (2002) Sui et al. (2002) Yu et al. (2002) Brummelkamp et al. (2002)
+1 3’ -200 -50 -25 Pol III DSE PSE A/Tbox TTTTTT snRNA +1 3’ -200 -45 Pol II 10 nt DSE PSE 3’box snRNA snRNA genes
Polylinker: BglII KpnI XhoI NheI BamHI NheI U1 prom psiUx (3100nt) sense antisense loop 3’box loop antisense sense U1 prom 3’box psiU shRNA Cloning in psiUx
psiUX BglII KpnI XhoI NheI BamHI NheI GGTACCCTCGAGGCTAGCGGATCCGCTAGC CCATGGGAGCTCCGATCGCCTAGGCGATCG prom snU1 pSP65 polylinker Digestion with Bgl II - Xho I The antisense sequence is specific for targeting lamin A mRNA Ligation In vitro annealed oligos 3'end +1 transcriptional terminator box 5'-GATCTCACGCAGCCACTTCGAGTACCCTTTGTGTAGGGGTACTCGAAGTGGCTGCGTTTACTTTCTGGAGTTTCAAAAGTAGAC-3' 3'-AGTGCGTCGGTGAAGCTCATGGGAAACACATCCCCATGAGCTTCACCGACGCAAATGAAAGACCTCAAAGTTTTCATCTGAGCT-5' sense 9 nt loop antisense Xho I Bgl II
Anti-lamin siRNAs GGGCAAUUGGCAGAUCAAGCGUU UUCCCGUUAACCGUCUAGUUCGC 5’- 3’- GGGCAAUUGGCAGAUCAAGCG UUCCCGUUAACCGUCUAGUUCGC GGGCAAUUGGCAGAUCAAGCG UUCCCGUUAAC UCUAGUUCGC gc Lamin A/C mRNA Lamin A/C mRNA control siRNAs GGGCAAUUGcgAGAUCAAGCGUU UUCCCGUUAACgcUCUAGUUCGC 5’- 3’-
A C * C U AUA AGGGCAAUUGGCAGAUCAAGCGUUGUG A GUCCCCUAUUCCCGUUAACCGUCUAGUUCGCAAGUAGACAC G 5’- psiUa-lam miR30 “loop” 3’- UG U UG * C U AUA AGGGCAAUUGGCAGAUCAAGCGUU GUCCCCUAUUCCCGUUAACCGUCUAGUUCGCGA 5’- psiUb-lam “9-base loop” 3’- UG U UG * 5’- GGGCAAUUGGCAGAUCAAGCGUU UUCCCGUUAACCGUCUAGUUCGCGA psiUc-lam “9-base loop” 3’- CG GC mut. GA GC A U * GGGCAAUUGGCAGAUCAAGCGUUUCUUCGCAUGAAUG G UUCCCGUUAACCGUCUAGUUCGCAAAGAAG UACUUAC U 5’- psiUd-lam “Rnt loop” 3’- U1-based anti-lamin A/C constructs
s + => as-loop B) psiU psiU A) U6 U6 mut mut NT NT c d a b d a b c 81 69 59 44 27 20 81 69 59 44 27 20 as-loop s as Probe a Probe a Probe amut D) C) Probe amut E) Probe U7 s-loop + => as
A C * C U AUA AGGGCAAUUGGCAGAUCAAGCGUUGUG A GUCCCCUAUUCCCGUUAACCGUCUAGUUCGCAAGUAGACAC G 5’- psiUa-lam miR30 “loop” 3’- UG U UG * C U AUA AGGGCAAUUGGCAGAUCAAGCGUU GUCCCCUAUUCCCGUUAACCGUCUAGUUCGCGA 5’- psiUb-lam “9-base loop” 3’- UG U UG * 5’- GGGCAAUUGGCAGAUCAAGCGUU UUCCCGUUAACCGUCUAGUUCGCGA psiUc-lam “9-base loop” 3’- CG GC mut. GA GC A U * GGGCAAUUGGCAGAUCAAGCGUUUCUUCGCAUGAAUG G UUCCCGUUAACCGUCUAGUUCGCAAAGAAG UACUUAC U 5’- psiUd-lam “Rnt loop” 3’- U1-based anti-lamin A/C constructs
Cell, 115, 199-208, 2003 Asymmetry in the Assembly of the RNAi Enzyme Complex D.S. Schwarz, G. Hutvágner, T. Du, Z. Xu, N. Aronin, and P.D. Zamore The siRNA strand whose 5’ is more easily unwound is preferentially kept.
UG U UG * C U 5’- psiUb-5a AUA AAACUUGAUCUGCCAAUUGCCCUU GUCCCCUAU UUUGAACUAGACGGUUAACGGGGA 3’- swap swap swap NT NT NT 5a 5a 5a b b b 26 - 26 - Probe a Probe a Probe U7 UG U UG UG U UG * * C U C U 5’- 5’- psiUb-lam psiUb-swap AUA ACGCUUGAUCUGCCAAUUGCCCUU GUCCCCUAU UGCGAACUAGACGGUUAACGGGGA AUA AGGGCAAUUGGCAGAUCAAGCGUU GUCCCCUAU UCCCGUUAACCGUCUAGUUCGCGA 3’- 3’- Asymmetric siRNA strand production
psiU A) U6 mut NT d a b c 81 69 59 44 27 20 psiU b a c NT U6 d mut Lamin A Lamin C GAPDH Probe a C) E) Probe amut % of Lamin A/C Probe U7 Activity of the siRNAs produced by psiUx
siUcmut-lam siUc-lam sense sense antisense antisense loop loop 3’box U1 prom U1 prom LTR GAG SV40 PURO LTR ** ** 3’box Selectable vectors for stable expression of siRNAs pBabe puro
Long-term expression and activity of the siRNAs A) pBabe-Uc pBabe-Ucmut 2 weeks 4 weeks 2 weeks 4 weeks 1 2 3 1 2 3 1 2 3 1 2 3 siRNAs Probes: a amut U2 snRNA B) pBabe-Ucmut pBabe-Uc 1 2 3 1 2 3 NT Lamin A Lamin C GAPDH
b-amyloid protein microglial cell RNAi IAA-94 CLIC1 activation TNFa e NO2- release Inflammation Neurotoxicity b-amyloid protein Toxicity neuron
700 UG U UG UG U UG UG U UG UG U UG * * * * C U C U C U C U 5’- 5’- 5’- 5’- psiUb-clic2 psiUb-clic4 psiUb-clic1 psiUb-clic3 AUA AGCGCCGAAGAUGAGGGCAUCUUU GUCCCCUAU UCGCGGCUUCUACUCCCGUAGAGA AUA ACUGCAACCUGCUGCCAAAGCUUU GUCCCCUAU UGACGUUGGACGACGGUUUCGAGA AUA ACGGGACUGGACAUAUUUGCAAUU GUCCCCUAU UGCCCUGACCUGUAUAAACGUUGA AUA AGAUGAUGAAGAGAUAGAGCUAUU GUCCCCUAU UCUACUACUUCUCUAUCUCGAUGA 3’- 3’- 3’- 3’- 600 500 TNF-a pg/ml 400 300 200 100 0 Psiux Vector only psiUb- clic2 psiUb- clic4 untreated Ab Novarino G, Fabrizi C, Tonini R, Denti MA, Malchiodi-Albedi F, Lauro GM, Sacchetti B, Paradisi S, Ferroni A, Curmi PM, Breit SN and Mazzanti M (2004) “Involvement of the intracellular ion channel, CLIC1, in microglia-mediated b-amyloid-induced neurotoxicity.” J. Neuroscience 24,5322-30
A system for stable expression of short interfering RNAs in mammalian cells. Brummelkamp TR, Bernards R, Agami R. Science 2002 249,550-553 p53, is a transcription factor which is stabilised by radiation and plays a crucial role in the maintenance of cell-cycle arrest in G1 after DNA damage.
A system for stable expression of short interfering RNAs in mammalian cells. Brummelkamp TR, Bernards R, Agami R. Science 2002 249,550-553 Co-transfection with pBabe-puro and selection with puromicyn
Viral delivery • Adenovirus and lentivirus • More effective delivery to broader range of cell types (incl. non-dividing) • Creation of stable lines (lentivirus) • Use in vivo
Stable suppression of tumorigenicity by virus-mediated RNA interference. Brummelkamp TR, Bernards R, Agami R. Cancer Cell 2002 Sep;2(3):243-7 • RAS genes are frequently mutated in cancer • Regulate proliferation, differentiation and cell survival • Point mutations block the oncogenic protein RAS in a persistently activated form binding GTP • Wild type protein is essential
Stable suppression of tumorigenicity by virus-mediated RNA interference. Brummelkamp TR, Bernards R, Agami R. Cancer Cell 2002 Sep;2(3):243-7 Human U2-OS cells
Stable suppression of tumorigenicity by virus-mediated RNA interference. Brummelkamp TR, Bernards R, Agami R. Cancer Cell 2002 Sep;2(3):243-7
Stable suppression of tumorigenicity by virus-mediated RNA interference. Brummelkamp TR, Bernards R, Agami R. Cancer Cell 2002 Sep;2(3):243-7
Specific inhibition of gene expression using a stably integrated, inducible small-interfering-RNA vector. Van de Wetering et al. EMBO Reports 2003 4, 609-615
Specific inhibition of gene expression using a stably integrated, inducible small-interfering-RNA vector. Van de Wetering et al. EMBO Reports 2003 4, 609-615 CRC cells: colorectal cancer
Specific inhibition of gene expression using a stably integrated, inducible small-interfering-RNA vector. Van de Wetering et al. EMBO Reports 2003 4, 609-615 Northern blot Western blot
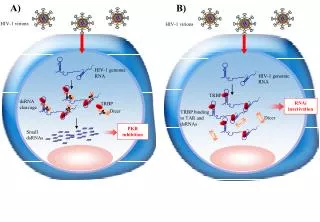
RNAi is a natural defense mechanism against viruses in plants but it is not known if it has a role in virus defense in mammals VIRAL TARGETS:essential genes for replication, genome regions in RNA viruses (HIV, HCV) CELLULAR TARGETS : host genes necessary for virus entry or for its life cycle, but not essential for cell survival Viruses as targets of RNAi
Inhibition of human immunodeficiency virus type1 replication in primary macrophages by using Tat- or CCR5-specific small interfering RNAs expressed from a lentivirus vector. Lee et al. Journal of Virology (2003) 77:11964-11972 Northern blot Western blot Virus replications as measured with p24
Inhibition of human immunodeficiency virus type1 replication in primary macrophages by using Tat- or CCR5-specific small interfering RNAs expressed from a lentivirus vector. Lee et al. Journal of Virology (2003) 77:11964-11972 Controllo 1 settimana 12 settimane The effect persists 12 weeks
Human Immunodeficiency Virus Type 1 Escape from RNA Interference. Boden et al. Journal of Virology (2003) 77:11531-11535 Inhibition effect is lost at 25 days, when an HIV variant appears, mutated in the middle of the target region
So, which is the way? • Target more than one gene at the same time! • Sinergistic effect • There should be no escape CCR5 Tat+CCR5 Tat CCR5 cellular target Tat viral target