Download
1 / 1
10 likes | 149 Views
Background. Rate & temperature curves. delay in neurophonic. delay in single units. Conclusions. Methods. Reversible inactivation of the ventral auditory brainstem by cooling. Eric Verschooten, Myles Mc Laughlin, Philip X. Joris

E N D
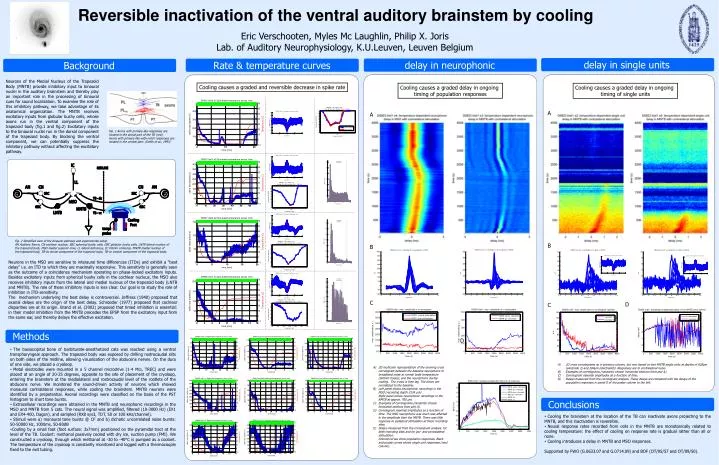
Background Rate & temperature curves delay in neurophonic delay in single units Conclusions Methods Reversible inactivation of the ventral auditory brainstem by cooling Eric Verschooten, Myles Mc Laughlin, Philip X. Joris Lab. of Auditory Neurophysiology, K.U.Leuven, Leuven Belgium Neurons of the Medial Nucleus of the Trapezoid Body (MNTB) provide inhibitory input to binaural nuclei in the auditory brainstem and thereby play an important role in the processing of binaural cues for sound localization. To examine the role of this inhibitory pathway, we take advantage of its anatomical organization. The MNTB receives excitatory inputs from globular bushy cells, whose axons run in the ventral component of the trapezoid body (fig.1 and fig.2) Excitatory inputs to the binaural nuclei run in the dorsal component of the trapezoid body. By blocking the ventral component, we can potentially suppress the inhibitory pathway without affecting the excitatory pathway. Cooling causes a graded and reversible decrease in spike rate Cooling causes a graded delay in ongoing timing of population responses Cooling causes a graded delay in ongoing timing of single units A A Fig. 1 Axons with primary-like responses are located in the dorsal part of the TB (red). Axons with primary-like-with-notch responses are located in the ventral part. (Smith et al., 1993) Fig. 2 Simplified view of the binaural pathway and experimental setup.AN Auditory Nerve, CN cochlear nucleus, SBC spherical bushy cells, GBC globular bushy cells, LNTB lateral nucleus of the trapezoid body, MSO medial superior olive, LL lateral lemniscus, IC inferior colliculus, MNTB medial nucleus of the trapezoid body, TB-dc dorsal component of the trapezoid body, TB-vc ventral component of the trapezoid body. B B • The basioccipital bone of barbiturate-anesthetized cats was reached using a ventral transpharyngeal approach. The trapezoid body was exposed by drilling rostrocaudal slits on both sides of the midline, allowing visualization of the abducens nerves. On the dura of one side, we placed a cryoloop. • Metal electrodes were mounted in a 5 channel microdrive (1-4 M, TREC) and were placed at an angle of 20-25 degrees, opposite to the site of placement of the cryoloop, entering the brainstem at the mediolateral and rostrocaudal level of the rootlets of the abducens nerve. We monitored the sound-driven activity of neurons which showed monaural contralateral responses, while cooling the brainstem. MNTB neurons were identified by a prepotential. Axonal recordings were classified on the basis of the PST histogram to short tone bursts. • Extracellular recordings were obtained in the MNTB and neurophonic recordings in the MSO and MNTB from 5 cats. The neural signal was amplified, filtered (10-3000 Hz) (EX1 and EX4-400, Dagan), and sampled (RX8 sys3, TDT, 50 or 100 kHz/channel). • Stimuli were a) monaural tone bursts @ CF and b) dichotic uncorrelated noise bursts: 50-50000 Hz, 1000ms, 50-80dB • Cooling by a small foot (foot surface: 3x7mm) positioned on the pyramidal tract at the level of the TB. Coolant: methanol passively cooled with dry ice, suction pump (FMI). We constructed a cryoloop, through which methanol at -30 to -40°C is pumped as a coolant. The temperature of the cryoloop is constantly monitored and logged with a thermocouple fixed to the exit tubing. Neurons in the MSO are sensitive to interaural time differences (ITDs) and exhibit a “best delay” i.e. an ITD to which they are maximally responsive. This sensitivity is generally seen as the outcome of a coincidence mechanism operating on phase-locked excitatory inputs. Besides excitatory inputs from spherical bushy cells in the cochlear nucleus, the MSO also receives inhibitory inputs from the lateral and medial nucleus of the trapezoid body (LNTB and MNTB). The role of these inhibitory inputs is less clear. Our goal is to study the role of inhibition in ITD-sensitivity. The mechanism underlying the best delay is controversial. Jeffress (1948) proposed that axonal delays are the origin of the best delay. Schroeder (1977) proposed that cochlear disparities are at its origin. Brand et al. (2002) proposed that timed inhibition is essential: in their model inhibition from the MNTB precedes the EPSP from the excitatory input from the same ear, and thereby delays the effective excitation. C D C D • 2D cross correlograms as in previous column, but now based on two MNTB single units at depths of 628µm (electrode 2) and 264µm (electrode5). Responses are to contralateral noise. • Examples of correlograms (randomly chosen horizontal sections from plot A). • Correlogram maximal amplitudes as a function of time. • Delays measured from the correlogram analysis. These delays are compared with the delays of the population responses in panel D of the poster column to the left. • 2D multicolor representation of the evolving cross correlogram between the baseline neurophonic to broadband noise at normal body temperature (bottom traces), and the neurophonic during cooling. The x-axis is time lag. The colors are normalized to the baseline. • Left panel shows neurophonic recordings in the MSO (recording depth 3134 µm)Right panel shows neurophonic recordings in the NMTB at approx. 700 µm. • Examples of correlograms (randomly chosen horizontal sections from plot A). • Correlogram maximal amplitudes as a function of time. The MSO neurophonic was much less affected in the amplitude than the MNTB. There was little response to ipsilateral stimulation at these recording sites. • Delays measured from the correlogram analysis, for both recording sites and for ipsi- and contralateral stimulation. Colored curves show population responses. Black and purple curves shows single unit responses (next column). • Cooling the brainstem at the location of the TB can inactivate axons projecting to the MNTB, and this inactivation is reversible. • Neural response rates recorded from cells in the MNTB are monotonically related to cooling temperature: the effect of cooling on response rate is gradual rather than all or none. • Cooling introduces a delay in MNTB and MSO responses. • Supported by FWO (G.0633.07 and G.0714.09) and BOF (OT/05/57 and OT/09/50).