Download

1 / 17
180 likes | 365 Views
This chapter introduces students to the development of the hominid line. It discusses the emergence of the Australopithecines and the evidence we have for their biology and lifeways. Early Hominids. Chronology of Hominid Evolution.

E N D
This chapter introduces students to the development of the hominid line. It discusses the emergence of the Australopithecines and the evidence we have for their biology and lifeways. Early Hominids
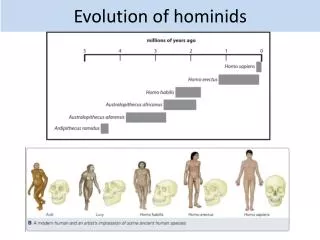
Chronology of Hominid Evolution • The Pleistocene (2 m.y.a. to 10,000 B.P.) is the epoch of human life. • Lower Pleistocene (2 to 1 m.y.a.): Australopithecus and early Homo • Middle Pleistocene (1 m.y.a. to 130,000 B.P.): Homo erectus and archaic Homo sapiens • Upper Pleistocene (130,000 to 10,000 B.P.): modern Homo sapiens • During the Pleistocene there were several ice ages, or glacials. • The glacials were separated by warm periods called interglacials. • The Würm was the last glacial (75,000 to 12,000 B.P.)
Ardipithecus • Ardipithecus fossils are generally considered ancestral to early Australopithecus and, ultimately, to Homo. • Ardipithecus fossils date to 5.8 – 4.4 m.y.a. • Presumably, Ardipithecus ramidus evolved into A. anamensis, a bipedal hominid from Kenya. • A. anamensis then evolved into A. afarensis, which is usually considered ancestral to all the later australopithecines.
Kenyanthropus • This species was identified in 1999 based on fossils from northern Kenya. • Kenyanthropus fossils date to 3.5 m.y.a. • Meave Leakey views Kenyanthropus as an entirely new branch to the early family tree.
The Five Species of Australopithecus • A. anamensis (4.2 m.y.a.). • A. afarensis (3.8? to 3.0 m.y.a.). • A. africanus (3.0? to 2.5? m.y.a.). • A. robustus (2.6 to 2.0 m.y.a.). • A. boisei (2.6? to 1.2 m.y.a.).
Australopithecus afarensis • Earliest definite A.afarensis remains are dated at 3.8 m.y.a. and (with Ardipithecus ramidus and A. anamensis) strongly support a very recent (8 m.y.a. at the most) divergence from common ancestry with chimpanzees, because of the clearly apelike features found in all three species. • Postcranial remains, particularly the pelvis, leg, feet, and spinal entry into the skull, all indicate bipedalism, and are thus clearly hominid. • A. anamensis remains (4.2 m.y.a.) also indicate bipedalism, and thus bipedalism predates A. afarensis.
Australopithecus afarensis (cont.) • Cranial remains show that afarensis was still remarkably “apelike” is some respects, in that the brain capacity was only slightly larger than a modern chimpanzee’s, and canines indicate considerable sexual dimorphism. • Fossils of molars and jaws indicate the beginnings of adaptation to a coarse (seeds, lots of sand) savanna diet. • A. afarensis young probably depended on their parents for a relatively long time.
Bipedalism • The most widely accepted theories stress the advantages provided by bipedalism in a habitat increasingly dominated by drier, savanna-like conditions. • The ability to see over long grass. • Bipedalism is more energy efficient and therefore advantageous in a grassland, where resources are more dispersed than in forests. • Bipedalism exposes less body surface area to solar radiation, which facilitates cooling and reduces moisture loss.
Gracile and Robust Australopithecines • There is some debate as to the nature of the distinction which existed between the gracile A. africanus and the robust A. robustus. • One model has africanus and robustus as separate species, whose life spans were at least partly contemporaneous. • Another model has africanus and robustus as sequential, with africanus being ancestral to robustus. • A third model has both groups as part of a single polytypic species, representing opposite extremes of variation within that species.
Gracile and Robust Australopithecines (cont.) • A similar debate obtains for A. robustus vs. A. boisei, but it is most likely that boisei developed from robustus into a separate, hyperrobust species, highly specialized (giant molars, sagittal crest, relatively small front teeth) for a savanna vegetation diet (cf., the discussion of the black skull, below). • Both africanus and robustus probably made tools from perishable material, which is therefore not preserved in the fossil record. • There was only slight increase in brain capacity throughout Australopithecus, but brain organization was basically human.
Homo Begins • Contemporaneous (2 m.y.a.) sets of teeth, very different in size, constitute the earliest evidence of a generic split: the larger set is attributed to A. boisei, the smaller to H. habilis, the first species of the genus Homo. • The distinctive early Homo trends are a rapid increase in brain size, increasingly elaborate toolmaking, and an increasing emphasis on hunting, but there remains considerable debate as to when and in what population these trends led to speciation (from an australopithecine to Homo habilis). • Johanson and White propose that A. afarensis effectively produced two populations, one of which evolved into the other australopithecines, the other evolving into Homo habilis.
The Black Skull • The 1985 discovery of the black skull (dated 2.5 m.y.a.), apparently an early A. robustus, has made for more possible models of the divergence between Homo and Australopithecus. • The black skull, so-called because of the coloration lent by the mineral content of the fossil itself, was a surprising mixture of australopithecine features, particularly given its relatively early date. • An apelike jaw and relatively small brain (primitive features). • A sagittal crest on the top of the skull (previously associated with the relatively modern hyperrobusts, thus surprising to be found on a skull so old).
Black Skull Interpretations • One interpretation: on one branch of the split produced by the afarensis population there was another split, with one branch leading to the gracile species (A. africanus), the other branch leading to black skull’s population (called A. aethiopicus), which in turn split to produce A. robustus on one branch and A. boisei on another. • Another interpretation: on one branch of the split leading from afarensis there is a second split, with one branch leading to A. robustus and A africanus, and the other leading to the black skull and, thence, to A. boisei.
KNM-ER 1470 • H. Habilis was transitional and co-existed with A. boisei. • In 1972, Richard Leakey found the skull KNM-ER 1470 at the site of Koobi Fora. • The skull exhibits a mixture of Homo and australopithecine features. • The cranial capacity is larger than the australopithecines (Homo). • The molars are similar to those of hyperrobust australopithecines. • Dating the skull has been problematic as some researchers date it to 1.8 m.y.a, while others believe it is older, dating to 2.4 m.y.a.
KNM-ER 1470 Interpretations • Some scholars believe that KNM-ER 1470 belongs to its own species, Homo rudolfensis, while others argue that KNM-ER 1470 belongs to H. habilis. • Some researchers believe KNM-ER 1470 lived earlier than H. habilis; others think that they lived at the same time. • Kottak argues that the only sure conclusion is that there were a suite of hominids in East Africa before and after the advent of Homo.
Toolmaking as a Competitive Advantage • The oldest known manufactured tools, dated at 2.6-2 m.y.a., were found in various parts of Africa and are grouped under the name Oldowan pebble tools, given them by the Leakeys in 1931. • There is some speculation that relatively advanced toolmaking by Homo ancestors might have created the environment which produced the generic Homo-Australopithecus split. • H. habilis may be as old as 2.4 m.y.a., which means it existed close to the time of the origin of stone tools. • Kottak argues, based on circumstantial evidence, it was likely that Australopithecines also made tools, if less well and of less permanent material than later Homo groups. • 1.6-1.8 m.y.a. saw a dramatic increase in the hunting efficacy of Homo, and during this period A. boisei may have been forced into an exclusively vegetarian niche, providing an example of competitive exclusion.
Australopithecus garhi and Early Stone Tools • In 1999, a new hominid species, A. garhi, was found in Ethiopia associated with stone tools and the remains of butchered animals. • This find is significant for three reasons. • It added a new species to the human family tree • It demonstrated that the thigh bone elongated one million years before the forearm shortened—to create current human proportions. • It showed that early stone tools were designed at getting meat and marrow from big game. • This is evidence for emergent cultural capacities, which grow exponentially with the appearance of Homo. • With increasing reliance on hunting, toolmaking, and cultural abilities, Homo became the most efficient exploiter of the savanna niche. • The last australopithecines may have been outcompeted and forced into ever more-marginal areas.