Download

1 / 46
560 likes | 1.18k Views
Motor Proteins. 2 Motor Systems. Actin-based motility: motor proteins are myosins Tubulin-based motility: motor proteins are kinesins and dyneins. Roles of the actin-based system. Cell Crawling - lymphocytes Growth cone extension

E N D
2 Motor Systems • Actin-based motility: motor proteins are myosins • Tubulin-based motility: motor proteins are kinesins and dyneins
Roles of the actin-based system • Cell Crawling - lymphocytes • Growth cone extension • ‘Muscle-like’ contraction of ovarian follicles, mammary gland ducts, etc. • Muscle contraction • Distribution of vesicles, intermediate filaments and organelles within the cytoplasm
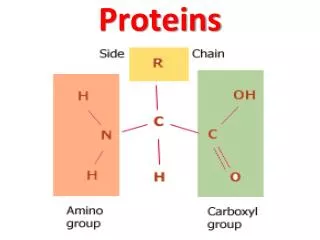
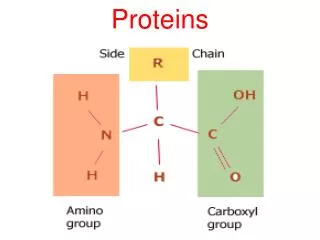
Actin-based motility: the Myosins The head is both actin-binding and ATP binding; the purple light chain has a regulatory role. Myosin II is muscle myosin.
There are lots of myosins • 18 different myosin families have been identified (I –XVIII) • There are a total of 40 myosin genes in the human genome – i.e. there are multiple copies of some of the genes
The Myosin Activity Cycle Myosin head energized, with bound ADP and Pi,, not attached to actin ATP hydrolyzed Myosin head attaches to actin HEAD DETACHED Head detaches from thin filament HEAD ATTACHED Head rotates - Powerstroke transmits force to thin filament – head is deenergized ATP replaces ADP and Pi on myosin head
Study of the movement of myosin against actin filaments – an in vitro motility assay • Preparation of actin cables pointing in same direction • Used cells from a giant alga that uses these cables for moving chloroplasts around. • Open algal cell and add yellow fluorescent beads coated with myosin, then add ATP and take time lapse photography
A series of exposures taken at intervals of 1s. This picture shows that the myosin coated beads moved along the actin cables Red dots are chloroplasts which fluoresce red
Speed of movement • Myosin coated beads moved unidirectionally and movement was dependent on ATP • The speed of beads coated with muscle myosin is 5 µm/s which is the same speed as the contraction of sarcomeres in muscle • Different myosins move at different speeds. Smooth muscle myosin moves at 1 µm/s.
Studies of movement due to a single myosin molecule • Use a setup in which focused laser beams create “optical traps”. These optical traps can hold small objects. The force is controlled by adjusting the intensity of the laser beam.
Actin filament is held in optical trap via one or two attached beads • Myosin concentrations are kept low so that only one myosin contacts the actin filament
ATP is also kept low so that only one ATP binds to each myosin head
Results show that the myosin pulls on the actin filament in a stepwise, or ratchet-like fashion displacement time
A single ATP molecule is hydrolyzed resulting in a power stroke and displacement of around 10 nm.
The force generated can also be determined and it is around 3-7 picoNewtons (pN)
Is the force and displacement what you would expect from the energy supplied by 1 molecule of ATP? DG= -12 kcal/mole for ATP hydrolysis = 16 x 10-21 cal per molecule ATP
1 pN x 10 nm displacement = 2.5 x 10-21 cal 3-7 pN of force generated ---> 7.5 to 17.5 x 10-21 calories. So the force and displacement for each step with actin/myosin motor is equivalent to the energy yield from the hydrolysis of 1 ATP
The neuron’s growth cone extends by building microfilaments at the + ends in response to growth cues from the environment…
Distribution of a vesicle along the actin network is polarized
Putting it together: Actin and Intermediate filaments • Myosin V is the link between actin, which serves as a rigid skeletal element, and the intermediate filament, which is being delivered to another part of the cell.
Roles of the tubulin-based system • Axoplasmic transport • Positioning organelles within the cell • Mitotic spindle • Cilia and flagella
Axoplasmic Transport • Anterograde – from cell body toward synapse – ie toward + end of the tubule - driven by kinesins • Retrograde – from synapse toward cell body – driven by dyneins • ‘Fast’ – works for cargo carried in vesicles – 50-400 mm/day. • ‘Slow’ – for individual protein molecules – net rate is less than 10 mm/day, but apparently this reflects a stop and go aspect of the process
Functions of axoplasmic transport • Delivers proteins, mitochondria, vesicles to synapses • Removes recycled proteins and organelles to cell body for destruction by lysosomes • Carries intracellular chemical messages from synapse to cell body • Delivers neuron-specific viruses (herpes and pox viruses) from peripheral sensory nerve endings to cell bodies in the CNS
Consequences of interfering with axoplasmic transport • Interruption of axoplasmic transport causes a traffic jam on the proximal side of the interruption and Wallerian degeneration of distal parts of neuron. • Anticancer drugs targeted against microtubules have neuronal toxicity.
Role of microtubules in positioning organelles within the cytoplasm 1. The endoplasmic reticulum is stretched towards the periphery by its connections to the microtubules. 2. Lysosomes are pushed toward the periphery by microtubules. 3. Three different kinesins are implicated in the movement of mitochondria along microtubule paths to the part of the cell where they are needed. 4. Dynein positions the Golgi; without microtubules, the Golgi breaks up into a lot of little vesicles that disperse in the cytoplasm. 5. Axoplasmic transport of vesicles to the axon terminals and relay of trophic substances (and herpes and chickenpox viruses) to the soma relies on the connections formed with kinesins and dyneins.
Microtubules are associated with motor proteins: Dynein and Kinesin (kinesin is thought to have evolved from myosin)
Microtubules are oriented: kinesin takes its cargo to the + end and dynein transports to the - end
Specific proteins mediate attachment of cargo to dyneins This cartoon is included to remind you that there must be a mechanism that designates particular vesicles for anterograde or retrograde transport
Some confusing terms • Organizing center for microtubules in an interphase cell = centrosome (usually); 1 centrosome consists of 2 centrioles; eachcentriole consists ofa pinwheel array of triplet microtubules. 1 basal body (cilia and flagella) = 1 centriole • Centromere = attachment point for spindle microtubule on chromosome = kinetochore
The sequence of mitotic processes • Nuclear membrane disassembled, chromosomes condense • Interphase microtubules dissassemble • Centrosome is duplicated – this initiates formation of mitotic spindle • At prometaphase, new microtubules form with their – ends attached to the centrosomes – the extending MTs randomly contact the kinetochores of the chromosomes and attach to them • During anaphase, sister chromosomes separate (anaphase A) and the spindle poles move further apart (anaphase B) • During telophase, daughter cells separate (cytokinesis- actin-myosin is involved in this) and the nuclear envelopes reform.
Some questions • How do chromosomes line up at the metaphase plate? • They are pushed there by net growth of microtubules, with polymerization occurring at the + end • How can microtubules draw sister chromosomes apart in anaphase A? • During anaphase A, spindle microtubules shrink by depolymerization near the + end - not the – end. No ATP is necessary for this process • How do the spindle poles move further apart from each other in anaphase B? • Dyneins push on microtubules in the overlap zone, while kinesins pull each end of the spindle toward the plasma membrane. ATP is required for this process
Polymerization pushes chromatids to the metaphase plate during prometaphase Depolymerization pulls chromosomes toward the spindle poles during anaphase A
The role of kinesins (green) and dyneins (pink) in anaphase B separation of spindle poles – at the same time, growth of microtubules at the + ends causes the spindle to elongate
Cilia and Flagella • Structure – basically the same structure • Differences: Cilia are shorter and numerous, whereas flagella are long and exist alone or as pairs. • The basal body that organizes the cilia or flagellum is identical in structure to the centrioles (right, below) that are present as a pair in the centrosome (left, below):
Functions of Cilia and Flagella • Cilia: • Respiratory airway (mucociliary escalator) • Oviduct (egg and sperm transport) • Flagella • Spermatozoa • Renal tubule
Basic facts about cilia and flagella • All eucaryotic cilia and flagella contain 9 outer bundles of doublet microtubules with a central singlet pair of microtubules – this entire structure is called an axoneme • Bending of the axoneme is the result of sliding of adjacent doublets relative to one another • Dynein arms generate the sliding forces – dyneins are attached to the b tubule of each doublet and their heads apply force to the adjacent a tubule
Cilia and Flagella: motility results from microtubule sliding within the axoneme
Cilia and Flagella: an axoneme (a cylinder of tubules: 9+2) connected to a basal body and covered by membrane
The axoneme The machinery inside cilia and flagella is constructed of a ring of 9 microtubule doublets (A=13, B=11) linked by nexin and powered by 2 dynein arms that have ATPase activity. The spokes link the ring to the inner 2 microtubules Inner arm dyneins are responsible for axoneme bending – outer arm dyneins just contribute to sliding but do not produce bends
Bacterial flagella and eucaryotic flagella are not homologous