Download

1 / 13
180 likes | 553 Views
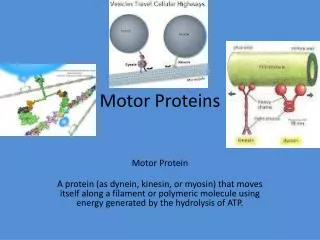
Motor Proteins - Introduction Part 1. Biochemistry 4000 Dr. Ute Kothe. Motor Proteins. Motor Proteins convert chemical energy into motion. chemical energy is derived from ATP hydrolysis motion is generated by conformational changes depending on the bound nucleotide.

E N D
Motor Proteins- Introduction Part 1 Biochemistry 4000 Dr. Ute Kothe
Motor Proteins • Motor Proteins convert chemical energy into motion. • chemical energy is derived from ATP hydrolysis • motion is generated by conformational changes depending on the • bound nucleotide Myosin Kinesin Dynein
Motor Protein Function Myosin (18 known classes, 40 different myosins in humans) • Movement along actin fibres • Muscle movement • Cytokinesis (cytoplasmic division, tightening of contractile ring) • Transport of cargo along microfilaments (vescicles etc.) Kinesin (16 classes) • Movement along microtubule tracks, usually to (+) end • Transport of cargos: vesicles, organelles, cytosolic components such as mRNAs & proteins, chromosomes Dynein (12 mammalian dyneins) • Movement along microtubule tracks,to (-) end, i.e. cell center • Cytoplasmic dyneins: transport of cargos such as vesicles • Axonemal dyneins: Movement of cilia and flagella
Tubulin • building block of microtubules • heterodimer of closely related Tubulin a & b • G proteins: N-terminal residues fold into G domain-like structure • a-tubulin’s GTP buried at subunit interface, nonexchangable, not hydrolyzed • b-tubulin’s GTP is solvent exposed until tubulin dimers polymerize • upon polymerization, a-tubulin from adjacent dimer provides catalytic residue to hydrolyze b-tubulin’s GTP; resulting GDP is nonexchangeable unless tubulin dissociates from microtubule Tubulin b + GDP Tubulin a + GTP Voet Fig. 35-89
Microtubules • Tubulins interact head to tail to form a long protofilament • Protofilaments align side by side in curved sheet • Sheet of 13 (9-16) protomers closes on itself to form microtubule • Microtubule lengthens by addition of tubulins to both ends (preferentially to + end, i.e. the end terminating in b-tubulins) Voet Fig. 35-92
Structure of the Axoneme • Bundle of microtubules called axoneme • coated by plasma membrane • Forms eukaroytic flagella & cilia Voet Fig. 35-102
Dynein • 1 or more heavy chains (motor domain) • several intermediate and light chains • motor domain is 7-membered ring, ATP-hydrolyzing unit • coiled-coil extension forms stalk that interacts via globular domain with microtubules • long stem (with intermediate and light chains) binds cargo Valle, Cell 2003; Voet Fig. 35-107
Conventional Kinesin • two identical heavy chains forming two large globular heads attaching to microtubules and a coiled-coil • two identical light chains interacting wit cargo • transports vesicles and organelles in (-) to (+) direction (towards cell periphery) Voet Fig. 35-94
Kinesin Structure • Globular head: tubulin-binding site & nucleotide binding site • flexible neck linker • a-helical stalk leading into coiled-coil • ATP hydrolysis triggers conformational change in neck linker via 2 switch regions: • When ATP is bound, neck linker docks with catalytic core • Upon ATP hydrolysis, the neck linker “unzips” Voet Fig. 35-95 & 96 & 97
Kinesin Cycle Voet Fig. 35-98
Hand-over-Hand Mechanism ATP-bound state: strong microtuble binding ADP-bound state: weak microtubule binding • ATP binds to leading head - globular kinesin head which is already bound to the microtubule and oriented towards (+) end • Neck linker of leading head “zips up” agains catalytic core • trailing head is thrown forward (trailing head has bound ADP and reduced affinity to microtubule): • Trailing head swings by ~ 160 Å, net movement of dimeric kinesin is ~ 80 Å = length of one microtubule dimer • ATP in new trailing head is hydrolyzed & phosphate released: affinity for microtuble decreases • ADP in new leading head dissociates • Two heads work in a coordinated fashion • ATP binding to leading head induces power stroke
Processivity • Kinesin is highly processive: it takes several 100 steps on a microtubule without detaching or sliding backwards! • How? • coordinated, but out of phase ATP cycle in both heads • one head is always firmly attached to microtubule Movie demonstrating kinesins processivity: http://www.proweb.org/kinesin/axonemeMTs.html
Dynein • 1 or more heavy chains (motor domain) • several intermediate and light chains • motor domain is 7-membered ring, ATP-hydrolyzing unit • coiled-coil extension forms stalk that interacts via globular domain with microtubules • long stem (with intermediate and light chains) binds cargo Valle, Cell 2003; Voet Fig. 35-107