Download
1 / 46
460 likes | 623 Views
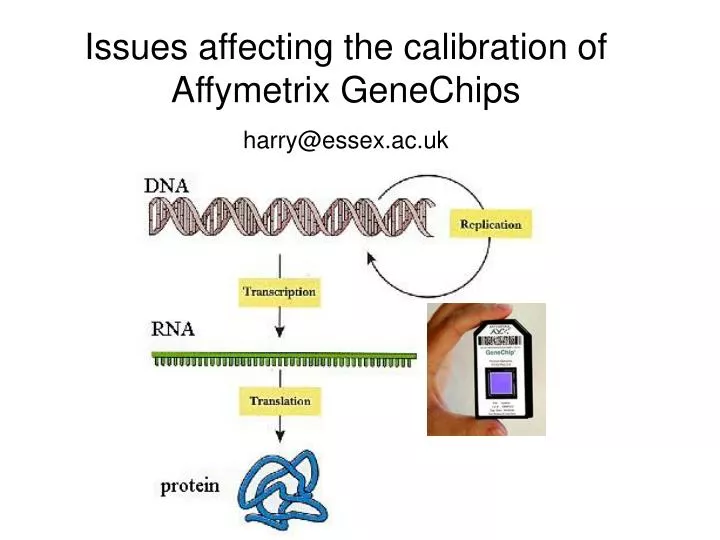
Issues affecting the calibration of Affymetrix GeneChips harry@essex.ac.uk. Overview of GeneChip technology and how gene expression is measured. How the data is calibrated. Biophysics of GeneChip technology. Biological issues such as splicing. Potential problems with parts of the data.

E N D
Issues affecting the calibration of Affymetrix GeneChips harry@essex.ac.uk
Overview of GeneChip technology and how gene expression is measured. How the data is calibrated Biophysics of GeneChip technology Biological issues such as splicing Potential problems with parts of the data
Many laboratories have an almost identical set-up for running GeneChips.
Probe cells of an AffymetrixGeneChip contain millions of identical 25-mers 25-mer
Detect fluorescence Remove partial hybrids by washing in a solution with a reduced salt content (phosphate backbones of nucleic acids have negative charge). Labelling with a fluorescent marker (on the Us). Hybridization Fragmentation of RNA to mean length of ~100 bases.
Reverse transcription has a chance of stopping at every base. This results in a population of aRNA copies which are biased towards the 3’ end.
Affymetrix microarrays 5’ 3’ GTGGGAATTGGGTCAGAAGGACTGTGGCTAGG GGAATTGGGTCAGAAGGACTGTGGC GGAATTGGGTCACAAGGACTGTGGC perfect match probe cells mismatch probe cells Probe-pairs scattered on chip
Probes can map to different transcripts. Such multiple targeting should be identified (Okoniewski and Miller BMC Bioinformatics 2006) Some probes no longer map to the most update transcript information for a gene. These probes will not detect expression related to the rest of the probeset to which they belong. There are a number of alternative Chip Definition Files, which group probes into probe-sets.
Probe cell (aka feature) Perfect Match (PM) Mismatch (MM) Probe pair Affymetrix probe set
Affymetrix software derives the intensity for each probe from the 75% quantile of the pixel values in each box.
Chip calibration Correct Background, Normalise, Correct for Cross Hybridisation, Expression Measure High-level analysis, biological interpretation
Background Fluorescence needs to be corrected e.g. MAS and RMA algorithms
Chips need to be normalised against each other. Each chip is a different colour e.g. invariant genes, lowess, quantiles
PA PB PC PD PE PA PB PC PD PE Chip 1 Chip 1 Chip 1 Chip 1 1 2 3 4 5 1.33 2.33 3.33 4.66 7 1 2 4 3 5 1.33 2.33 4.66 3.33 7 Chip 2 Chip 2 Chip 2 Chip 2 1 2 3 5 7 7 2.33 4.66 3.33 1.33 7 2 5 3 1 1.33 2.33 3.33 4.66 7 Chip 3 Chip 3 Chip 3 Chip 3 4.66 2.33 3.33 1.33 7 1.33 2.33 3.33 4.66 7 2 3 4 5 9 5 3 4 2 9 Order by ranks Average the intensities at each rank Reorder by probe RMA uses Quantile normalisation at the probe level
Expression Measure The intensities of the multiple probes within a probeset are combined into ONE measure of expression
Cross Hybridisation MAS 5.0 (Affymetrix) corrects for cross-hybridisation by subtracting the MisMatch signal from the Perfect-Match. RMA ignore the mismatches because they hybridise to the Perfect Signal.
MAS 5.0 (Signal) takes the Tukey bi-weighted mean of the difference in logs of PM and MM.
1-9 are different chips. dChip and RMA ‘model’ the systematic hybridisation patterns when calibrating an expression measure.
Spike-in measurements show there remains considerable signal at low concentrations.
Cross Hybridisation MAS 5.0 (Affymetrix) corrects for cross-hybridisation by subtracting the MisMatch signal from the Perfect-Match. RMA ignore the mismatches because they hybridise to the Perfect Signal. How can you measure cross-hybridisation without using the MisMatch signal? There is a need for a model of the physics of hybridisation (Naef and Magnasco 2003)
Size is important e.g. perfect match #13 = A, so mismatch #13 is T, and the complementary base in mRNA is also T/U T Pyrimidines (C & T) are small There will be no steric hindrance between the pyrimidine in the mismatch and the pyrimidine in the mRNA of interest. C Purines (G & A) are large G There will be a large steric hindrance between the purine in the mismatch and the purine in the mRNA of interest. A
GC content is important AT bonds have two hydrogen bonds. GC have 3 hydrogen bonds
Probes are ssDNA Target is RNA
Biotin labelling interferes with the hybridisation C & T (pyrimidines) are labelled. So GC* binds less strongly than CG, and AT* binding is weaker than TA. If the probe contains no C & T, it will hybridise well but with no fluorescence. If you have all C & T, it will have difficulty hybridising.
Van der Waals interactions between adjacent bases H-bond interactions between adjacent bases Nearest-neighbour interactions predict duplex kinetics and so sequence order is important (Santa Lucia) The binding energy of GAC is not the same as CAG
Wu and Irizarry (2004) have written GCRMA (which is available in Bioconductor). Lots of close sequences will hybridise to a given probe. Wu and Irizarry model the variation in hybridisation of these similar processes using a statistical model. GCRMA determines the contribution to the PM from Signal and from Non-Specific Hybridisation
GCRMA produces a good linear relationship between intensity and concentration
Once chips have gone through the DATCELExpression Measure process, changes in gene expression between conditions or over time can be observed. m=log2(Fold Change), a=log2(Average Intensity) The change in expression between two conditions for all the genes on an array can be viewed on a MA plot
Some genes are represented by multiple probe-sets. Probe-set A Probe-set B If they are measuring the same gene the signals should be up and down regulated together! Is that always true? No Stalteri and Harrison, 2007, BMC Bioinformatics, 8:13
Alternative Splicing results in different permutations of exons from the same gene. Sometimes introns are also included. Each of these permutations results in a different protein. Alternative Splicing affects > 50% of our genes. It is the most obvious way in which 25,000 genes in the Human Genome can produce ~100,000 proteins.
Probes map to different exons. Because of alternative splicing, some of the exons may be upregulated whereas others may be downregulated.
Single poly(A) site Alternative poly(A) sites in the 3’-mostexon Alternative poly(A) sites in differentexons Tian et al. (2000) Alternative Polyadenylation
Probes map to different sides of a polyadenylation signal. Because of alternative polyadenylation, some of the probes may be upregulated whereas others may be downregulated.
Comparing probes with runs of Gs. Number of contiguous Gs Mean Correlation 3 0.14 4 0.42 5 0.49 6 0.62 7 0.75 We are only looking at a small fraction of the entire probe, yet it is dominating the effects across all experiments.
A tetrad of Guanines can bind to each other through Hoogsteen Hydrogen bonds with the help of a central cation. G-quadruplexes are prevalent in telomeres (single stranded DNA at the end of chromosomes). G-quadruplexes are thermally stable. G-quadruplexes take a range of topologies.
Adjacent probes within a cell on a GeneChip have the same sequence – a run of Guanines will result in closely packed DNA with just the right properties to form quadruplexes. Upton et al. 2008 BMC Genomics, 9, 613
Parallel G-quadruplexes have a left-handed helical twist. We suggest 4 probes can efficiently form a “Maypole”. Outside the corset of the “G-spot”, the probes have little affinity for bases of the same sequence and the phosphate backbones will repel each other. Inside the G-spot the bases are on the inside and cannot bind target. GGGG
Hybridization kf Probe + Target Duplex kr Dissociation R is the Gas Constant, and T is temperature. G = - RT ln K All spontaneous physical and chemical changes take place in the direction of a decrease in free energy, G < 0
Phosphates on chains of nucleic acids have a negative charge. There is a coulomb block of hybridization on microarrays (Vainrub and Pettitt 2002). The environment caused by probe-probe interactions acts to modify the hybridization of RNA. Hagan and Chakraborty 2004, Journal of Chemical Physics The strength of binding depends upon probe density G = - RT ln K
GGGG GGGG GGGG G = - RT ln K Probes that are not bound in G-quadruplexes will have a reduced probe density in the immediate environment of the runs of Guanines. This will result in very effective nucleation, and binding, with respect to hybridization to the rest of the probe. The binding will efficiently occur in the G-spot. Any RNA molecule with a run of Cs will hybridize. Thus, there will be enhanced correlations between all the probes that are able to form G-quadruplexes.
Kerkhoven et al. 2008, PLoS ONE 3(4): e1980 Probes containing GCCTCCC will hybridize to the primer spacer sequence that is attached to all aRNA prior to hybridization.
GeneChips require calibration in order to reliably combine the information from multiple probes. Biophysics of GeneChip technology has been studied in a lot of detail – but there is still things to learn. Algorithms such as GCRMA attempt to provide a physics-based transformation between light and RNA concentration. Biological issues such as splicing Potential problems with particular sequences
Bioinformatix, Genomix, Mathematix, Physix, Statistix, Transcriptomix are needed in order to extract reliable information from Affymetrix GeneChips Thank you for your attention.




















