Download
1 / 45
450 likes | 632 Views
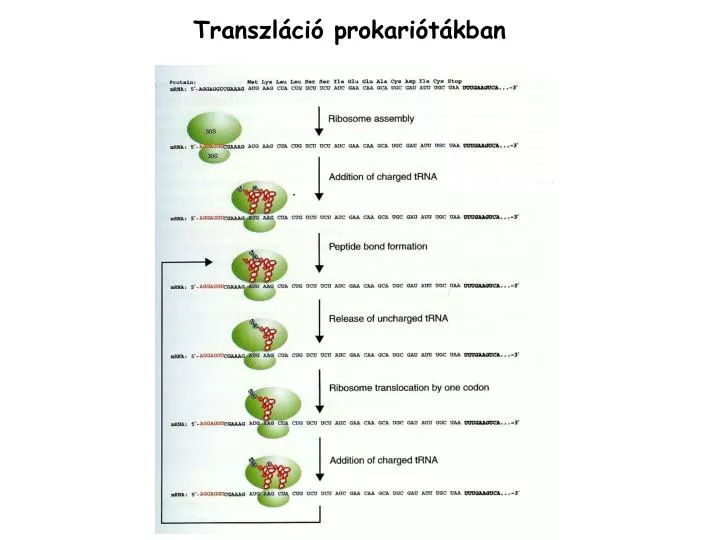
Transzl áció prokariótákban. 30S. 30S *. ( 30S + IF1, IF2, IF3, fMet-tRNS Met ). k 1. ( preiniciációs komplex). 30S * + mRNA. PK. k -1. k 2. ( iniciációs komplex). PK. IK. k -2. k 3. ( transzlációs komplex). IK. TK. A transzláció hatékonyságát meghatározó elemek.

E N D
30S 30S* ( 30S + IF1, IF2, IF3, fMet-tRNSMet) k1 ( preiniciációs komplex) 30S* + mRNA PK k-1 k2 ( iniciációs komplex) PK IK k-2 k3 ( transzlációs komplex) IK TK A transzláció hatékonyságát meghatározó elemek K1 nagy, ha az SD és a rRNS közti kölcsönhatás erős. k2 a SD és a transzlációs start kodon közti távolság, és a régió másodlagos szerkezetének függvénye.
A transzláció hatékonyságát meghatározó elemektechnikai megközelítés
gyakoriság kodon AUG 91 % GUG 8 % UUG 1 % Shine - Dalgarno szekvencia GGAGG a tökéletes komplementer a 16S rRNS 3'végéhez, pl. GAAG GGAG mutáció (maltóz operon) 10 x növekedés a termék mennyiségében a SD szekvencia hosszának növelése nem feltétlenül növeli a transzlációt Hasonlóan a promóterekhez, túl erős kötődés a SD szekvencia és a 16S rRNS 3’ vége között a transzlációt gátolhatja. Transzlációs start kodon
Az AUG előtti szekvenciák 20x-os különbség UAU, CUU>>UUC, UCA, AGG ("-1 triplet) G-k kerülendők Az AUG utáni szekvenciák 30x -oskülönbségek lehetnek AAA(Lys) > AAG(Lys) > UUU(Phe), AUC/A(Val), GUA(Ala)... A természetes gének között is az AAA(Lys) és GCU(Ala) transzlációs szabályozás a 2. kodonnal +10 régió AT gazdag
Távolság a start kodon és a SD között - meglehetősen érzékeny, általban 4 - 13 bázispár megengedett. - Génfüggő, egy gén esetén általában csak kicsit változhat. A köztes szekvencia összetétele “A” jó, ha van (2x-es stimulus), “C” mindegy “U” a "-4"-es pozícióban hatékony “G” gátol (3x-os gátlás) Upstream nemtranszlálódó szekvencia Nem nagyon vizsgált, de kell, mert eltávolítása megszünteti a transzlációt (ld lac). Minimum 10 bp kell. Esetleg valamilyen faktorok kötődnek hozzá.
Kodon felhasználási preferencia Bár a kodonok gyakorlatilag univerzálisak, a különböző sejtféleségek különböző gyakorisággal használják az azonos aminosavakat kódoló tripleteket. Ha a termeltetendő fehérje génjének kodon használata nagyon eltér a gazdasejtétől, akkor meg kell szintetizáltatni a gént a gazda sejt preferenciájának megfelelően. Alternatív megoldás: a gazdasejt kodon preferenciájának megváltoztatása, ritka kodonokhoz tartozó tRNS gének bevitelével
Sphingomonas paucimobilis AmAcid Codon Number /1000 Fraction Gly GGG 208.00 14.99 0.17 Gly GGA 70.00 5.04 0.06 Gly GGT 117.00 8.43 0.10 Gly GGC 827.00 59.59 0.68 Glu GAG 476.00 34.30 0.60 Glu GAA 318.00 22.91 0.40 Asp GAT 297.00 21.40 0.35 Asp GAC 558.00 40.20 0.65 Val GTG 386.00 2 7.81 0.43 Val GTA 40.00 2.88 0.04 Val GTT 65.00 4.68 0.07 Val GTC 412.00 29.69 0.46 Ala GCG 630.00 45.39 0.40 Ala GCA 142.00 10.23 0.09 Ala GCT 108.00 7.78 0.07 Ala GCC 687.00 49.50 0.44 Met ATG 376.00 27.09 1.00 Arg AGG 35.00 2.52 0.04 Ile ATA 19.00 1.37 0.03 Arg AGA 12.00 0.86 0.01 Ile ATT 117.00 8.43 0.17 Ser AGT 24.00 1.73 0.04 Ile ATC 570.00 41.07 0.81 Ser AGC 232.00 16.72 0.3 Thr ACG 228.00 16.43 0.33 Lys AAG 465.00 33.50 0.86 Thr ACA 29.00 2.09 0.04 Lys AAA 76.00 5.48 0.14 Thr ACT 41.00 2.95 0.06 Thr A CC 383.00 27.60 0.56 Asn AAT 151.00 10.88 0.37 Asn AAC 252.00 18.16 0.63 Trp TGG 250.00 18.01 1.00 End TGA 37.00 2.67 0.74 Cys TGT 21.00 1.51 0.11 Cys TGC 167.00 12.03 0.89 End TAG 5.00 0.36 0.10 End TAA 8.00 0.58 0.16 Tyr TAT 213.00 15.35 0.56 Tyr TAC 170.00 12.25 0.44 Leu TTG 71.00 5.12 0.06 Leu TTA 4.00 0.29 0.00 Phe TTT 8 6.00 6.20 0.14 Phe TTC 542.00 39.05 0.86 Ser TCG 204.00 14.70 0.31 Kodon felhasználási preferencia – táblázat [gbbct]: 50 CDS's (13879 codons) egy adott gén esetén a kodon preferencia alapján a gén E. coli-ban, vagy élesztőben való expresszálhatósága in silico becsülhető.
STOP kodon UAA UGA UAG kötődő faktor RF1, RF2 RF1 RF2 STOP kodon UAA >> UGA, UAG Az esetek 80 %-ában: UAAU
KÉT CISZTRONOS EXPRESSZIÓS RENDSZER 1 Cisztronos rendszer SD 5’ 3’ ATG struktúrgén STOP mRNS 2 Cisztronos rendszer termeltetendő fehérje 1. SD 1. STOP 5’ 3’ ATG 1. cisztron ATG 2. cisztron mRNS 2. SD 2. STOP Az 1. cisztron tetszőlegesen tervezhetú, de: - ne legyen túl hosszú, hogy ne terhelje feleslegesen a sejtet (felesleges műtermék)néhány aminosav - AT gazdag legyen a másodlagos struktúrák elkerülése végett.
1 Cisztronos rendszer SD 5’ 3’ ATG struktúrgén STOP mRNS 2 Cisztronos rendszer termeltetendő fehérje 1. SD 1. STOP 5’ 3’ ATG 1. cisztron ATG 2. cisztron mRNS 2. SD 2. STOP Az 1. cisztron STOP kodon pozíciója a 2. cisztron Shine-Dalgarno szekvenciájához képest meghatározó a transzláció hatékonysága szempontjából. PÉLDA bGH (marha növekedési hormon) Egy cisztronos rendszerben < 0.4 % fehérje Két cisztronos rendszerben 1. cisztron ...GGAGGAATAACATATG...2. cisztron. elrendeződésben 24 % fehérje termeltethető
Kifordított – antiriboszómális kötőhelyen alapuló - rendszer
PROTEOLÍZIS Abnormális fehérjék proteolitikus lebontási mechanizmusa E. coli-ban Abnormális fehérjék, sejten belüli aggregátumok ATP függő endoproteázok polipeptidek < 1500 Da endoproteázok aminosavak peptidek oldékony mono-, di- és tripeptidázok
La (Lon) a fő ATP függő proteáz • 87 kDa egységekből felépülő homotetramer, • ATP és Mg2+ függő, az ATP-áz aktivitás nem a polipeptid kötésfelbontásához, hanem a degradálandó fehérjén való végiglovagláshoz kell, • in vitro 2 ATP/ peptidkötés, in vivo ez változhat, és ennél nagyobb. • hasonló fehérjék előfordulnak magasabbrendűekben is, pl. egy májmitondriális proteáz immunológiailag keresztreaktív • abnormális fehérjék, és rövid élettartamú (pl. szabályozó fehérjék bontása) • szerin proteáz, hidrofób jellegű szekvenciáknál hasít • alapállapotban inaktív, a szubsztrát fehérje hozzákapcsolódása aktiválja, • a lon gén terméke, és a sokk fehérjékhez kapcsolódó szabályozás alatt áll: 32 faktor, ami a htpR (rpoH) gén terméke
Proteolitikus ciklus inaktív proteáz (4 ADP) protein szubsztrát 4 ADP 4 PO43- 4 Mg 2+ 4 ATP - Mg2+ alloszterikus aktiválás aktív proteáz (4 ATP)
Genetikai vonatkozások léteznek inszerciós, deléciós lon mutánsok A lon- mutáció másodlagos hatásai Poliszaharid burok képzés (főleg alacsonyabb hőmérsékleten, mint 34oC) Megoldás: • 37oC-on növeszteni • galE struktúrgének inaktiválása • cps (capsule) struktúrgének inaktiválása • rcsB, rcsA szabályozó régió mutánsok A lon mutánsok UV érzékenyek DNS sérülés filamentáció Gazdag médiumon akár letális is lehet ok: SOS indukálható sejtosztódás inhibitor SulA. Megoldás: - ne használjunk yeast extraktot, hanem tryptont -sulA mutánsok
Tulajdonság Ti (Clp) proteáz ClpP ClpA aktivitás fehérje degradáció ATP hidrolízis mellett peptidáz ATP független ATP-áz fehérje aktivált Kofaktor Mg2+ - Mg2+ Inhibítor MalNEt, iPr2P-F iPr2P-F MalNEt Stabilizálás ATP, glicerin hőstabil ATP, glicerin natív méret 700 kDa 260 kDa 160 kDa alegységek 6 ClpA + 12 ClpP 21 kDa 81 kDa Ti (Clp) a másik ATP függő proteáz • 81 (ClpA) és 21 kDa (ClpP) alegységekből álló heterodimer, ez multimerizálódik, a natív enzim kb 700 kDa móltömegű • ATP és Mg2+ faktorok szükségesek hidrofób részeknél hasít, de a specificitás eltér a La-tól • az alegységek külön multimerizálódnak • HtpR szabályozás alatt áll
Genetikai vonatkozások • Rendelkezésre állnak clp mutánsok • a mutáns egyedül nem, de a lon- nal együtt hatékony Egyéb proteázok • OmpT külső membránhoz kapcsolódó proteáz • DegP periplazmatikus proteáz • T4 fág fertőzés stabilizálja a fehérjéket • pin gén terméke: La inhibitor
FÚZIÓS FEHÉRJÉK A fúziót gén szinten hozzuk létre úgy, hogy a fúziós partnerek génjeit a megfelelő leolvasási keretben úgy klónozzuk össze, hogy az 5’ vég felőli génhez tartozó transzlációs stop kodon ne szerepeljen. Így egy polipeptidláncot kapunk, amelyben a kiinduási fehérjék egymást követőrégiókként helyezkednek el. ELŐNYEI 1. Kis peptidek esetén a fúzió proteolitikus stabilitást jelenthet 2. Óriási előny a tisztítás során, affinitás oszlopok 3. szignálpeptiddel való fúzió, szekretált fehérje (limitált) 4. Az in vitro hasítás sokszor pontosabb, intaktabb N-terminális szekvenciát eredményez
Fúziós fehérjék oldhatósága Oldhatatlan "iclusion bodies" trpEII, vagy cII, nincs proteolízis, differenciális centrifugálással az "inclusion body" könnyen tisztítható, probléma: nem aktív fehérje pl sometostatin, inzulin A, B, ellenanyag termelés Oldható forma biológiailag aktív fehérje, affinitás oszlopon való tisztítás problémák a stabilitással, kevésbé jósolható
Egyéb hasznosítási lehetőségek 1. a szignál peptidekkel való fúzió előnyei (ld. később) a periplazmában, illetve az extracelluláris térben oxidatívabb környezet, diszulfid hidak kialakulása, pl. EGF, IGF A periplazmában az összfehérje 4%-a található, értelemszerűenkevesebb proteáz,könnyebb tisztítás 2. A fúzionált fehérje linkerként szolgál, és így a kívánt fehérje immobilizálható 3. funkcionális, struktúrális vizsgálatok, pl. immunológiai felhasználás alkalmasan választott fúziós partner esetén nincs szükség hasításra. 4. megfelelő hasítás után az intakt fehérje tetszőleges biokémiai, biofizikai vizsgálata
fúziós partner célfehérje I. 1. affinitás kromatográfia III. 2. affinitás kromatográfia II. hasítás tiszta fehérje A fúziós fehérjék tisztításának elve
P P P T T T P T S X A X X B A fúziós fehérjék típusai I. I. Szekréció Megjegyzés pl. humán növekedési hormon alkalikus foszfatáz szignál szekvenciával II. Polimerizáció fehérje-élettartam növekedés, pl. proinzulin 1-2 percről 60 perc, IGF 200x-os növekedés X X III. C-terminális fúzió a fúziós partner intakt szabályozó régiója használható, pontosabb proteolitikus hasítás IV. N-terminális fúzió amino terminális szekvenciák meghatározása, a proteolititkus hasítás csonkot hagyhat a “B” N-terminálisán
P P P P P T T T T T S A X A X B X A B S X I A fúziós fehérjék típusai II. V. A szekréció és a C-terminális fúzió kombinációja a termék folyamatosan kinyerhető affinitáskromatográfiával VI. Kettős fúzió nagy tisztaságú, nagy stabilitású termék, hátrány: két tisztítás, hasítás szükséges A B X VII. Szekréció-inszerció membrán fehérjék termeltetése a membránban
Fehérje Származási hely molekulasúly (kDa) szekréció ligand b-galaktozidáz Escherichia coli 116 - TPEG, APTG Protein A Staphylococcus aureus 31 + IgG Z szintetikus 7 + IgG CAT Escherichia coli 24 + klóramfenikol sztreptavidin Streptomyces 13 + biotin PhoS Escherichia coli 36 + hidroxilapatit Protein G Streptococci 28 + albumin MBP Escherichia coli 40 + keményítô GST Escherichia coli 26 - glutation poliarginin szintetikus 1-3 - ioncsere poliglutamát szintetikus 1-2 - ioncsere polihisztidin szintetikus 1-7 + Ni2+, Zn2+, Cu2+ Fúziós partnerfehérjék CAT: klóramfenikol-acetiltranszferáz; Z: A Protein A fehérje IgG kötô doménje; PhoS: foszfát kötô fehérje; MBP: maltóz-kötô fehérje; GST: glutation S-transzferáz; TPEG: p-aminofenil--d-tiogalaktozidáz; APTG: p-aminofenil--d-tiogalaktozid.
hasító ágens hasítási hely brómcián - Met hangyasav - Asp Pro - hidroxilamin - Asn Gly - FÚZIÓS FEHÉRJÉK HASÍTÁSI MÓDOZATAI I. KÉMIAI ENZIMATIKUS aminopeptidázok 1 - 3 aminosavat hasítanakaz N-terminálisról karboxipeptidázok 1 - 2 aminosavat hasítanak a C-terminálisról
ENZIM HASÍTÁSI HELY kollagenáz - Pro - Val Gly - Pro - enterokináz - Asp - Asp - Asp - Lys Xa faktor - Ile - Glu - Gly - Arg trombin - Gly - Pro - Arg tripszin - Arg, Lys klosztripain - Arg Ala64-szubtilizin - Gly - Ala - His - Arg FÚZIÓS FEHÉRJÉK HASÍTÁSI MÓDOZATAI II. ENZIMATIKUS Endopeptidázok
A fehérje szekréció Nómenklatúra: szekréció: az extracelluláris térbe export: az sejtmembránokba vagy a periplazmatikus térbe Előnyei • A tisztítás sokkal egyszerűbb, esetleg folyamatos üzemű lehet • Az periplazma illetve az extracelluláris tér sokkal oxidatívabb, korrektebb protein folding • A fehérje degradáció sokkal csekélyebb mértékű, proteolitikusanstabilabb termékek
SZEKRÉCIÓ E. coli-ban • nem hatékony vagy nem teljes transzport a membránon keresztül • alacsony transzport-kapacitás, stresszválasz, idő előtti fehérjedegradáció • ha a transzporthoz a fehérje poszttranszlációs módosítása szükséges,akkor ennek hiánya kis molekulákra, mint növekedési faktorok vannak jó eredmények nagy molekulák esetén problémák lehetnek, pl az ER hiánya (módosítások színhelye) vannak fehérjék, amelyek nem szekretálhatóak, esetleg letális, pl -galaktozidáz fúziós fehérjék exportja
Fő protein transzport mechanizmusok bacteriumokban Kotranszlációs transzport SRP, signal recognition particle, és receptoraE. coli homológ: Ffh forty five homolog, FtsY Poszttranszlációs transzport Sec útvonal: egyszerű kofaktor mentes polipeptidekre Tat útvonal: több alegységes enzimekre, amelyek kofaktorokat is tartalmaz(hat)nak transzport: a fehérje összeszerelődése után
Sec útvonalE. coli-ban • - szignál szekvencia, • - szignál peptidázok, szignál peptidázI (lep), Ala-X-Ala felismerőhely • - szignál peptidázII (lsp), szignál szekvenciák eltávolítása lipoproteinekrôl • - sec A, B, D, E, Y gének mutációja a prekurzorok felhalmozódásához vezet, • Némi átfedés van közöttük. • - Régebben prl nómenklatúra is volt • SecA, 102 kDa, citoplazmatikus, és belső membránkötött, ATP-áz aktivitás, • kölcsönhatás a szállítandó fehérjével • SecB szekréció kompetens konformációért felelős • SecYEFG a csatorna kialakításáért felelős • SecD a periplazmatikus térbe való “leválásért”
hasítóhely hasítóhely “C” “N” “H” “C” “N” “H” A V S fordulat P G A G S A G hidrofób hélix, sok A és L, nem lehet P, K, R, D, E, H, R hidrofób hélix, sok A és L, nem lehet P, K, R, D, E, H, R L 1 K vagy R semleges vagy negatív 1 K vagy R semleges vagy negatív COOH COOH -1 +1 -3 -1 +1 fM fM G fordulat D lipoprotein kiválasztó szignál A X SZIGNÁL SZEKVENCIÁK GRAM- BAKTÉRIUMOKBAN I. Standard szignál peptid Lipoprotein szignál peptid
SZIGNÁL SZEKVENCIÁK II. Gram-pozítív baktériumok • a szignál peptid szignifikánsan hosszabb • kiterjedtebb “H” régió Eukarióták • a “H” régióban a Leu dominál • nem olyan szigorú a (-1, -3) szabály • a hasítóhely után nincs kitüntetett aminosav az eukarióta szekretált fehérjéket az E. coli export mechanizmusafelismer(het)i, illetve fordítva
INICIÁCIÓ TRANSZLOKÁCIÓ KISZABADULÁS periplazma H+ H+ D Y E Y E Y E SPáz G G G A A A ATP ATP B ADP+Pi ADP+Pi citoplazma C N citoplazma N+ C+ Átjutás a C-terminális véggel, ez az áltatános Átjutás az N-terminális véggel, inszerció A Sec függő fehérje transzlokáció modellje E. coli-ban
A prokarióta és eukarióta transzportmechanizmusokösszehasonlítása
Az SRP (Ffh-FtsY) útvonal membrán fehérjékre jellemző A Sec és az SRP közötti választás: “trigger” faktor
erősen hidrofób + N Signal sequences for targeting Sec szignál peptid n-region h-region c-region SRP: erősen hidrofób szignál, nem hasad le Twin-arginine szignál peptid moderáltan hidrofób + N S/T-R-R-x-F-L-K n-régió h-régió c-régió
A periplazmitikus hidrogenázokbioszintézisecitoplazmatikusösszeszerelődési modell Holoenzim Periplazma csatorna Citoplazma szignál peptid kofaktor(ok) prekurzor
periplazma N N N membrán TatB TatE C C TatA C TatC N C citoplazma The twin-arginine translocase (Tat) proteins
membrán TatC TatB TatA citoplazma aktív formát felvettenzim A TAT modell




