Download
1 / 20
200 likes | 399 Views
MITOCHONDRIÁLNÍ PROTEÁZY. Lukáš Stibůrek. INTRACELULÁRNÍ PROTEOLÝZA. Koncept „proteinového obratu“ – 40. léta minulého století Rudolf Schoenheimer na Columbia Univ – společně s Davidem Rittenbergem pokusy s 15 N-značenými AK

E N D
MITOCHONDRIÁLNÍ PROTEÁZY Lukáš Stibůrek
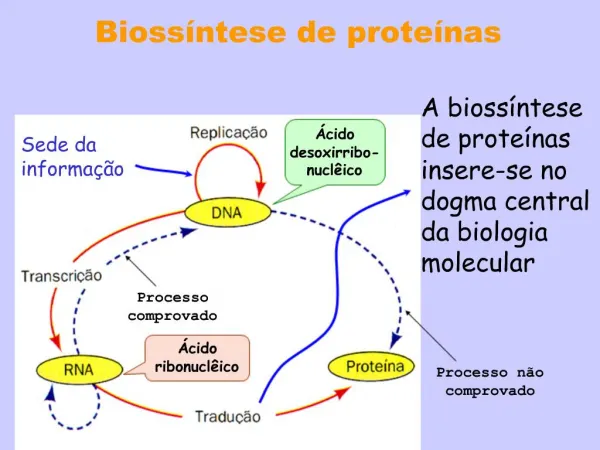
INTRACELULÁRNÍ PROTEOLÝZA Koncept „proteinového obratu“ – 40. léta minulého století Rudolf Schoenheimer na Columbia Univ – společně s Davidem Rittenbergem pokusy s 15N-značenými AK Lysozom – objeven v polovině 50. let Christian de Duve ATP-dependentní proteolýza za nepřítomnosti lysozomů (retikulocyty) – 1977 Alfred Goldberg Ubikvitin-dependentní proteolýza – 1980 Aaron Ciechanover et al. Institute of Technology, Haifa 2004 Nobelovacena – Aaron Ciechanover, Avram Hershko, Irwin Rose
MITOCHONDRIÁLNÍ PROTEOLÝZA • A. NESELEKTIVNÍ – MAKROAUTOFAGIE (Lysozom) • B. SELEKTIVNÍ – • – VNĚJŠÍ MEMBRÁNA – 26S Proteazom • – MEZIMEMBRÁNOVÝ PROSTOR, VNITŘNÍ MEMBRÁNA A MATRIX – Mitochondriální proteázy • selektivní intramitochondriální proteolýza - různé poločasy života mitochondriálních proteinů • ~5% mitochondriálních proteinů degradováno za hodinu – vysoká stabilita • 5-10% nově importovaných preproteinů degradováno – efektivita mitochondriální biogeneze >90%
MITOCHONDRIÁLNÍ PROTEÁZY (funkční třídy) • 1. Procesující peptidázy – odštěpení peptidů signalizujících mitochondriální lokalizaci • 2. ATP-dependentní proteázy – kontrola proteinové kvality, proteolytická maturace mito proteinů • 3. Oligopeptidázy– štěpení intramitochondriálních peptidů na aminokyseliny
PROCESUJÍCÍ PEPTIDÁZY Mitochondriální procesující peptidáza (MPP) – konzervovaná heterodimerní metalopeptidáza (a,b podjednotky) Mitochondriální intermediátní peptidáza (MIP) – monomerní metalopeptidáza štěpící oktapeptid z preproteinů po jejich štěpení MPP Peptidáza vnitřní membrány (IMP) – heterotrimerní komplex ukotvený ve vnitřní membráně s aktivní doménou exponovanou do IMS Atp23 (KUB3) – konzervovaná metalopeptidáza se specifickou funkcí – štěpí presekvenci mtDNA kódované podjednotky Atp6 (yeast). Bifunkční charakter – dále funguje jako šaperon Atp6 při její asemblaci do komplexu ATP syntázy. Rhomboidní proteáza (Pcp1/PARL) – serinová proteáza s katalytickou doménou uvnitř svých hydrofóbních membránových segmentů, štěpí transmembránové proteiny přímo v membráně
ATP-DEPENDENTNÍ PROTEÁZY – odstranění nadbytečných, poškozených a špatně sbalených polypeptidů, proteolytická maturace importovaných polypeptidů ClpXP proteáza – heterooligomerní komplex lokalizovaný v matrix skládající se z ClpP podjednotek se serin-peptidázovou aktivitou a ClpX šaperonových podjednotek zajišťujících rozpoznání a rekrutování substrátů – zevrubně charakterizovaná u prokaryot – proteolytická ClpP podjednotka chybí u kvasinek LON proteáza – homooligomerní serin-proteázový komplex lokalizovaný v mitochondriální matrix – afinita k mtDNA – její podjednotky podstupují proteolytickou maturaci
ATP-DEPENDENTNÍ PROTEÁZY AAA PROTEÁZY Obsahují Walker A a B motivy v tzv. AAA doméně zajišťující vazbu a hydrolýzu ATP, dále pak proteolytickou doménu s konzervovanou HEXGH sekvencí i-AAA proteáza – homooligomerní komplex podjednotek YME1 zakotvený ve vnitřní membráně s aktivní doménou v IMS m-AAA proteáza – heterooligomerní komplex podjednotek Spg7 a Afg3L2 zakotvený ve vnitřní membráně s aktivní doménou v matrix SPG7 – hereditární spastická paraplegie AFG3L2 – hereditární ataxie SCA28
OLIGOPEPTIDÁZY • velmi málo charakterizované • delece genů pro mitochondriální oligopeptidázy nezpůsobuje • kvasinkám žádnou popř. větší újmu: • – funkční redundance jednotlivých enzymů • – existence ABC transporteru Mdl1 který zajišťuje export • nadbytečných peptidů z matrix • – spekuluje se o signalizační funkci těchto peptidů (retrográdní signál) • Thimet oligopeptidáza (Prd1/saccharolysin) – • degradace peptidů produkovaných i-AAA proteázou • Mop112/PreP olygopeptidáza – • degradace peptidů produkovaných i-AAA proteázou • Bleomycin hydroláza Lap3 – • duální lokalizace (cytosol/matrix)
YME1L je mitochondriální integrální membránový protein s C-koncem exponovaným do IMS, nativní molekulovou hmotností 600-1100 kDa a markantními rozdíly v tkáňové expresi
Stabilní RNAi YME1L v HEK293 buňkách vede k efektivnímu snížení hladin YME1L mRNA
Stabilní RNAi YME1L vede k efektivnímu snížení hladin YME1L proteinu, akumulaci Ndufb6 a Cox4 podjednotek a změně paternu fragmentů OPA1
Stabilní RNAi YME1L vede k zpomalení proliferace takových buněk a k fragmentaci a zeslabení mitochondriální sítě
Stabilní RNAi YME1L vede k abnormální morfogenezi mitochondriálních krist, po reexpresi YME1L-FLAG dochází k reverzi tohoto fenotypu
Stabilní RNAi YME1L vede k akumulaci subkomplexů komplexu I a IV OXPHOS a snížené respirační aktivitě komplexu I
Ndufb6, Cox4 a Cox2 jsou membránově lokalizovány a koimunoprecipitují s YME1L E543Q z YME1L RNAi buněk
Reexprese YME1L-FLAG v YME1L RNAi buňkách vede k supresi akumulace Ndufb6 a Cox4, exprese Ndufb6-FLAG v HEK293 buňkách vede k indukci tvorby subkomplexů komplexu I
Stabilní RNAi YME1L vede ke zvýšení citlivosti k oxidativnímu poškození (H2O2, 200 μM ) mitochondriálních membránových proteinů (DNP, 2,4-dinitrofenyl)
Stabilní RNAi YME1L vede ke snížené rezistenci k staurosporinem (STS, 2μM ) a H2O2 (200 μM)indukované programované buněčné smrti (apoptóze)
Koimunoprecipitace radioaktivně (35S) značených mitochondriálních translačních produktů s YME1L E543Q a jejich zvýšená stabilita v YME1L buňkách
PODĚKOVÁNÍ Mgr. Jana Česneková Mgr. Daniela Fornůsková, PhD MUDr. Laszló Wenchich, PhD Mgr. Olga Brantová, PhD MUDr. Josef Houštěk, DrSc. Prof. MUDr. Jiří Zeman, DrSc.