Download
1 / 28
350 likes | 689 Views
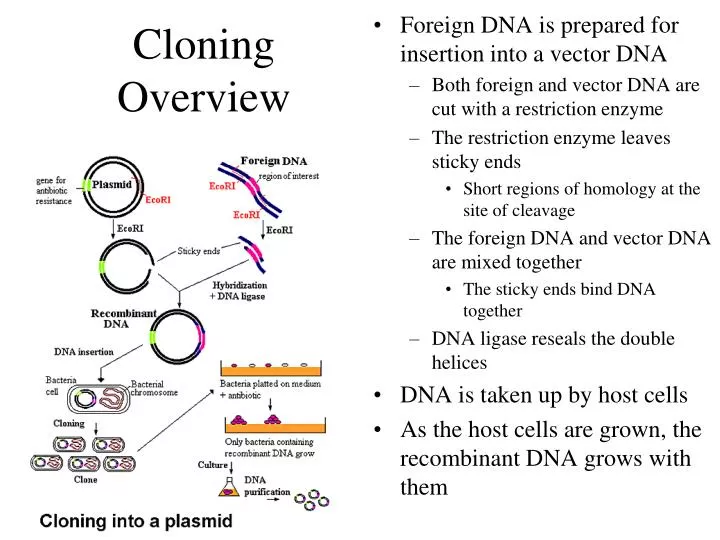
Cloning Overview. Foreign DNA is prepared for insertion into a vector DNA Both foreign and vector DNA are cut with a restriction enzyme The restriction enzyme leaves sticky ends Short regions of homology at the site of cleavage The foreign DNA and vector DNA are mixed together

E N D
Cloning Overview • Foreign DNA is prepared for insertion into a vector DNA • Both foreign and vector DNA are cut with a restriction enzyme • The restriction enzyme leaves sticky ends • Short regions of homology at the site of cleavage • The foreign DNA and vector DNA are mixed together • The sticky ends bind DNA together • DNA ligase reseals the double helices • DNA is taken up by host cells • As the host cells are grown, the recombinant DNA grows with them
Libraries • Libraries are made when DNA is cloned indiscriminately • All possible DNA from a foreign source is inserted into vectors one fragment at a time • Bacteria take up the vectors en masse one vector per bacterium • Propagation of the bacteria creates a culture that contains all of the DNA from a foreign source, fragmented into small pieces, with each piece in its own bacterial clone
Three general types of libraries • Conceptually represent DNA, RNA and protein libraries • Genomic libraries • This is used to clone genes • cDNA libraries • Made from mRNA, this contains sequence representing message • Expression libraries • A cDNA library that expresses the foreign proteins
Genomic Library • A genomic library contains all of the chromosomal DNA of a cell • DNA is purified and fragmented into pieces ranging from a few thousand bases to hundreds of thousands • The size of the fragments depends on the capacity of the vector to contain and propagate the DNA • The fragments are ligated into a vector and the vector propagated in a suitable host cell culture • Each piece of chromosomal DNA is then grown within a foreign host cell
cDNA Libraries • A cDNA library contains the sequences of (primarily) mRNA found in a cell • These sequences are propagated following conversion of a single stranded RNA molecule into double stranded DNA through the action of reverse transcriptase • They lack the transcriptional control and intronic sequences found in genomic clones • They are useful • In understanding structure and function of an mRNA • For example, the nucleotide sequence of an mRNA defines the exons of a gene • In expressing eukaryotic proteins in bacteria
Expression libraries • These are cDNA libraries in a special form of vector that permits transcription of the incorporated cDNA • Proteins or protein fragments then appear with bacterial host cells that are normally not present • These can be identified based on their antigenicity or activities • By this route a cDNA sequence can be isolated based on identification of its protein product • Proteins can also be made in quantity and purified more easily when made in bacteria
Why clone? • Analytical purposes • The DNA sequence and the structure/function relationships of a gene can be determined in isolation from the surrounding DNA of the genome by purifying the gene through cloning • A single gene is lost in the background of the genome • Cloning it isolates it. • Practical purposes • An isolated gene can be • Expressed to produce a protein in vivo or in vitro • Commercial purposes • Therapeutic purposes • Manipulated to change its sequence • This makes new genes with original proteins and properties • Cloning creates an unlimited supply of identical copies
Cloning reagents • Enzymes and buffers • Restriction enzymes • DNA ligase • And sometimes (for cDNA clones) • Reverse transcriptase and its allied reagents • Vectors • Host cells • And • Microbiological supplies • Radioactive or fluorescence labeled nucleotides
DNA ligase • While we have looked at ligase as an enzyme used during DNA replication, DNA ligase used during cloning procedures comes from the bacterial virus T4 • The enzyme is fragile and reactions are carried out at low temperature • It is capable of resealing the cut ends of restricted DNA, including blunt ends • Typically DNA is not purified away from restriction enzymes, but simply heated following restriction to denature the restriction enzyme • Then ligation is not defeated by recutting
Reverse transcriptase • This is used for converting an RNA transcript into DNA such that it can be cloned. • This is called copy DNA or cDNA • mRNA is primed with oligo dT for reverse transcription • RT makes a single stranded DNA copy of the RNA • The RNA is completely degraded and several strategies are employed to prime second strand synthesis • A hairpin loop naturally forms • But this loses sequence on the 5’ end • Oligo dA or dG can be enzymatically polymerized from the 3’ end • Then oligo dT or dC can be used as with the first strand
The hairpin loop and ragged ends of the new duplex cDNA are digested with a single strand specific nuclease making the duplex blunt ended • Then “linker” DNA is ligated onto the duplex • Linker DNA is a palindromic duplex oligonucleotide with a blunt end recognized by a restriction enzyme • Here it is HindIII • The linker DNA is then digested (with Hind III in this example) and the resulting DNA represents • The mRNA • A Hind III sticky end • This can be ligated into a vector as though it were genomic DNA that had simply been fragmented by Hind III
Vectors • DNA with an independent origin of replication and some selectable or differential markers • A selectable marker permits a host cell to survive in otherwise lethal environments • A differential, non-selectable marker permits identification of a bacterium by its appearance • Plasmids, viruses (and viral derivatives) and artificial chromosomes • All forms of cloning are technical variations on plasmid cloning
PBR322 • This is an artificial plasmid vector that has educational value, but is rarely used anymore • Commercial plasmid vectors are more versatile • PBR322 has two genes for antibiotic resistance • Amp and tet • There is a single site for the restriction enzyme Bam H1 in the tet gene • Inserting foreign DNA into this site inactivates the gene
The host • May be prokaryotic or eukaryotic • Must take up recombined DNA • Technical approaches to introducing DNA into bacterial cells • Add DNA to cells directly • Transformation • Bacteria take up plasmids • Transfection • Bacteria take up viral vectors • Eukaryotes take up any vectors • Electroporation • Drive DNA into cells with electric field • Must support growth of the recombined DNA
Colony screens • Once recombinant DNA has been taken up by a host, a successful transformation needs to be identified • Bacterial colonies are grown on a nutrient plate • If the foreign DNA was purified before it was inserted into the vector, then the selectable markers provide enough information to identify successful clones • However if the foreign DNA was not purified, then the bacterial colonies may overlayed with a membrane and lysed in situ on the membrane • The bacteria and DNA will stick to the membrane • Successful clones hybridize to radioactive complementary DNA like a southern blot
What to do with the DNA? • Once recombinant DNA is in a host, the host can be grown and plasmid easily isolated • Sequence it • This can be done directly on purified recombinant DNA • Express it • Proteins may be made in quantity • Mutate it • Site specific mutations are possible • Once a sequence is known, it is possible to alter any nucleotide by design • New proteins may be designed • Control elements of the inserted DNA may be studied and altered
Examples of medical relevance • Insulin Dependent Diabetes Mellitus (diabetes type I) • This results from an autoimmune destruction of the pancreatic beta cells • The body can no longer make insulin • Therapy requires monitoring of blood sugar and administration of insulin depending on glucose levels • Insulin originally came from animal sources • This molecule eventually elicited an immune response • Cloning technology made it theoretically possible to clone human insulin • There would be no immune response to this
Cloning insulin • The amino acid sequence of insulin was known, so a synthetic DNA sequence (probe) complementary to the insulin gene was available • The gene sequence is not exactly predictable due to the degeneracy of the code, but close enough to insure a unique identification • The conditions at which probe is washed off of a membrane are made less harsh (lowering the stringency of the wash) • This allowed imperfect hybrids of a sufficiently long probe to survive the wash
cDNA clones were necessary • The insulin gene has two introns, one of which interrupts the coding sequence of the gene • In order to express the protein, a cDNA clone was needed that eliminated the intron • Bacteria could not process mRNA from a cloned eukaryotic gene • mRNA from an insulinoma (pancreatic beta cell tumor) was isolated and cDNA made • This represented every message in the cell • Insulin mRNA represented a fraction of the total • All of the cDNA was inserted into plasmids at once • It wasn’t possible to purify insulin mRNA first • Each vector got a cDNA from a random mRNA from the insulinoma
Identifying the insulin cDNA clone • The culture of bacteria transformed with the insulinoma cDNA’s is called a library • It represents all of the mRNA in the cell • Bacteria were plated and the colonies “lifted” onto a membrane • Probing the membrane with a synthetic complementary sequence identified colonies that contained the insulin cDNA
Expressing the clone • Once the cDNA was isolated, the “insert” was removed from the vector and cloned into a vector that contained control sequences for RNA polymerase • The control sequences were the lac promoter • Transcription of the insulin gene could be increased with IPTG • Bacteria were transformed with this recombinant vector and insulin protein was synthesized from the transcript polymerized by RNA polymerase
Problems with expressing eukaryotic genes in a bacterium • But the insulin could not be properly processed in bacteria • The signal peptide and the center peptide could not be removed by the bacterium • It was also difficult to purify because the signal peptide caused aggregation of the protein within the bacterium
Another approach • The A and B chains represented individual polypeptides that are normally produced by processing of preproinsulin • It was necessary to remove the signal peptide from the clone prior to expression • The gene was cloned as a fusion protein with beta galactosidase • The gene, lacking the signal sequence but containing an N terminal methionine was used to replace the 3’ end of the betagalactosidase gene • This meant that expression of the cloned fusion gene produced a protein that was betagalactosidase on its amino terminal end and proinsulin (with an extra methionine) on the carboxyterminal end • This helped in purification because beta galactosidase did not aggregate and was easily purified • But the insulin part had to be separated from beta galactosidase
Reconstitution of insulin from individual chains • The extra methionine is specifically recognized and cleaved by the chemical reagent cyanogen bromide • This splits the insulin away from the beta galactosidase • Proinsulin was then mixed with C peptidases, that are made in pancreatic beta cells • This freed the A and B chains (now linked through disulfide bridges) producing insulin
Present day • The complete cDNA (including signal peptide) have been cloned into yeast • The yeast contain ER and signal peptidase, and they have also been engineered to contain the protease that cleaves the internal c-peptide form insulin • The yeast secrete human insulin • This circumvents the costly procedures necessary to purify insulin away from bacteria and then from the beta-gal.
Gene therapy for IDDM type I? • In immunodeficient mice, IDDM I can be cured with pancreatic beta cell transplantation • But beta cells are killed by an autoimmune response in people • Transplanted beta cells would simply be killed by the immune system • The use of insulin as a drug could be circumvented by putting an active insulin gene inside a patient • What kinds of problems might you expect in attempting this? • How would you control the gene? • In what cell type would you put it? • What other systems in the host cell would be required for proper expression of insulin?
Beta cell regulation • The pancreatic beta cell responds to elevated blood glucose by releasing stored insulin through exocytosis • Properly regulated insulin requires • Targeting to exocytic vesicles • This is due to recognition of preproinsulin structure within the ER and Golgi • Recognition systems must be present • The signal peptide and C-peptide must be removed • Storage prior to release • Exocytic vesicles must form containing mature insulin • They must be sequestered until a signal for release is received • Responsiveness of exocytosis to blood glucose • Exocytosis involves elevated calcium levels that promote fusion of the exocytic vesicle with the plasma membrane
Other human diseases potentially amenable to gene therapy • Most active • Severe combined immune deficiency (SCID) • Especially deficiency of adenosine deaminase • The expression of the gene needn’t be controlled and is expressed in rapidly growing cells (stem cells of the hematopoietic system) • Can’t find or transfect stem cells? • Cystic fibrosis • Lack of a chloride channel • Also expression needn’t be controlled • Accessible target cells (alveolar cells create the main clinical problem) • Delivery systems inadequate or unstable DNA? • Probability for a therapy to work increases if the expression levels of the protein don’t matter and that the defect is due to a missing enzyme