Download
1 / 43
510 likes | 1.15k Views
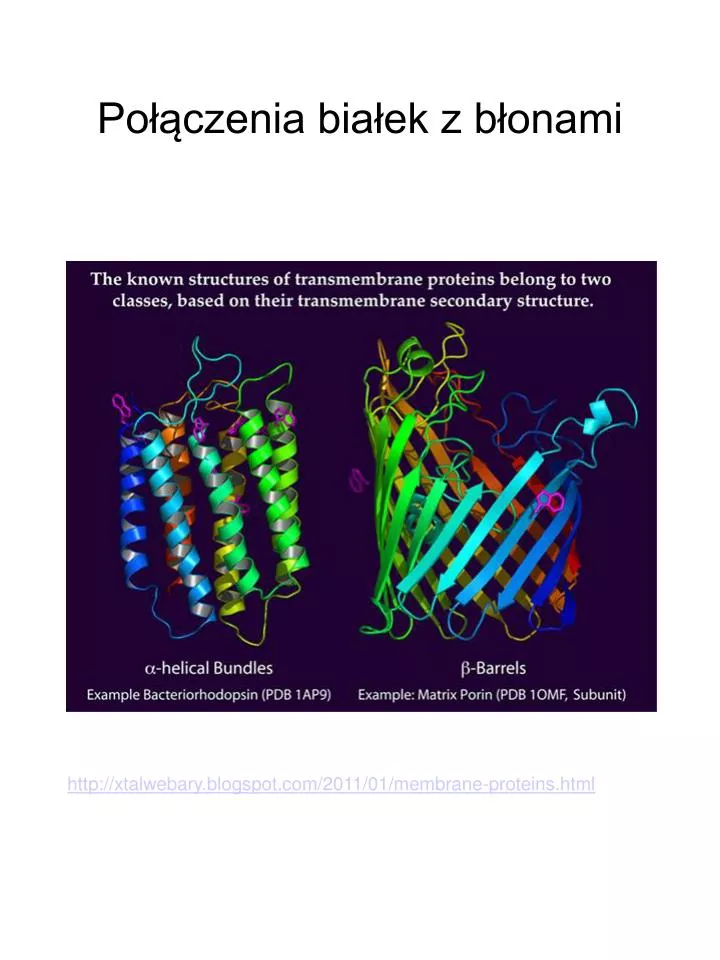
Połączenia białek z błonami. http://xtalwebary.blogspot.com/2011/01/membrane-proteins.html. inozytol N-acetyloglukozoamina mannoza galaktoza etanoloamina. GPI - glikozylofosfatydyloinozytol. Białka zakotwiczone przez GPI znajdują się na zewnątrz komórki.

E N D
Połączenia białek z błonami http://xtalwebary.blogspot.com/2011/01/membrane-proteins.html
inozytol N-acetyloglukozoamina mannoza galaktoza etanoloamina GPI - glikozylofosfatydyloinozytol
Białka zakotwiczone przez GPI znajdują się na zewnątrz komórki http://www.bio.cam.ac.uk/~dupree/dupree_gpi.jpg
Przykłady białek zakotwiczonych w błonie przez GPI • Białka powierzchni leukocytów • CD16b – jeden z receptorów Fc IgG • CD14 – receptor LPS • CD52 – małe białko (61 aa) o niewyjaśnionej funkcji • Białka adhezyjne • jedna z form N-CAM (neural cell adhesion • molecule) • Enzymy • PH20 – hialuronidaza plemników • MT4-MMP i MT6-MMP (membrane type • matrix metalloproteinases) • anhydraza węglanowa 4 (kardiomiocyty, • śródbłonek) • Białka powierzchni pierwotniaków • białka Toxoplasma, Leishmania • Inne: białko prionowe PrP • Białka z kotwicą GPI są uwalniane z błony przez fosfolipazę C lub fosfolipazę D aktywowane przez wiele bodźców.
Białko prionowe zakotwiczone w błonie przez GPI http://www.nature.com/nature/journal/v412/n6848/fig_tab/412739a0_F4.html
ZAKOTWICZANIE BIAŁEK W BŁONACH • WEWNĄTRZ KOMÓRKI • Mirystylacja przy N-końcu • Palmitylacja reszt cysteiny • Farnezylacja przy C-końcu • Geranylogeranylacja przy C-końcu
MIRYSTYLACJA • (kwas mirystynowy: C14, nasycony) • do mirystylacji wykorzystywany jest mirystylo-CoA • mirystylacja zachodzi kotranslacyjnie (są wyjątki) • mirystylacji ulegają białka o sekwencji N-końca: Met-Gly (Met-Gly-X-X-X-Ser/Thr) • Metionina jest najpierw usuwana przez aminopeptydazę metionylową, a następnie dochodzi do mirystylacji reszty glicyny przez enzym:transferazę N-mirystylową (NMT) Wright MH et al, J Chem Biol (2010) 3:19–35
Przykłady białek ulegających mirystylacji: • kinaza białkowa A, • podjednostki alfa białek G, • białka z rodziny Src (transdukcja sygnału), • oksydoreduktaza NADH:ubichinon, • białka wielu wirusów • Znaczenie mirystylacji: • umożliwia odwracalne zakotwiczanie białek w • błonach • współuczestniczy w tworzeniu trzeciorzędowej • struktury białek – stabilizuje konformację • tworzy miejsce oddziaływania z innymi białkami Silne wiązanie do błony wymaga oprócz mirystylacji drugiego efektora: sekwencji aminokwasów zasadowych lub palmitylacji
Zakotwiczenie białek w błonie przez kotwicę mirystylową jest procesem odwracalnym: Wright MH et al, J Chem Biol (2010) 3:19–35
PRZEŁĄCZNIK MIRYSTYLOWY Wiązanie poprzez resztę mirystylową do błony może podlegać regulacji poprzez mechanizm przełącznika mirystylowego. in out Przykład: rekoweryna – białko siatkówki oka regulujące aktywność rodopsyny. Przy podniesionym stężeniu wapnia – konformacja „out”’ przy normalnym poziomie wapnia konformacja „in”.
Inny sposób regulacji: zniesienie oddziaływania aminokwasów zasadowych z błoną przez zmianę ładunku białka wywołaną fosforylacją Białka MARCKS - myristoylated alanine-rich C-kinase substrate W formie nieufosforylowanej oddziałują z filamentami aktynowymi. Biorą udział w procesie sekrecji, egzocytozy, transportu, wpływają na kształt komórek i ich ruchliwość. Fosforylacja (przez PKC) – zwiększenie ładunku ujemnego – odłączenie od błony.
Potranslacyjnamirystylacja Proces odkryty podczas badań nad apoptozą Jednym z etapów apoptozy jest utrata funkcjonalności mitochondriów W procesie apoptozy nie tylko Bid ulega mirystylacji. Znane są jeszcze inne, cztery zidentyfikowano. Wright MH et al, J Chem Biol (2010) 3:19–35
PALMITYLACJA • kwas palmitynowy (C16, nasycony) • Do palmitylacji wykorzystywany jest • palmitylo-CoA, reakcję prowadzą enzymy • błonowe PAT (transferazy palmitylowe) • Palmitylacja zachodzi potranslacyjnie • Palmitylacji ulegają reszty cysteiny – • tworzy się wiązanie tioestrowe Istnieje wiele enzymów PAT (23 u ssaków) o rozmaitej lokalizacji wewnątrzkomórkowej (aparat Golgiego, błona ER, błona komórkowa, endosomy) i pewnej specyficzności.
SVM – synaptic vesicle membrane PM – plasma membrane PAT – transferaza palmitynowa APT – tioesteraza acyl:białko S. Baekkskov, J. Kanaani, Molecular Membrane Biology, 2009; 26: 4254
Palmitylacji ulegają cztery grupy białek: • białka transmembranowe – palmitylacja dotyczy reszt cysteiny znajdujących się jeszcze w obrębie błony lub tuż poza nią • (np. TGF-a, receptor transferyny, białka wirusowe, receptory o 7 domenach transmembranowych); palmitylacja jednej lub kilku reszt cysteiny. • białka prenylowane (białka Ras) • białka mirystylowane (często sekwencja: Met-Gly-Cys) • inne białka By: Charollais et al. Molecular Membrane Biology, 2009, 26, 55-66
Reszta palmitylowa może zostać odłączana od białek przez specyficzną tioesterazę: APT (acyl:protein thioesterase). PALMITYLACJA (jako jedyna spośród procesów przyłączania kotwic) jest procesem ODWRACALNYM Czy na pewno? – Nie…. Istnieją doniesienia o możliwości istnienia demirystylacji
Jakie elementy dotyczące funkcjonowania białek mogą zmieniać się pod wpływem palmitylacji? • Lokalizacja: • błona komórkowa (receptor transferyny) • receptor mannozo-6-fosforanu (RMP) • mikrodomeny błonowe (tetraspaniny, CD4, • integryny beta4 • Oddziaływanie z innymi białkami (integryna beta4, podjednostka receptora AMPA, RMP) • Przekaz sygnału (jeden z receptorów serotoninowych) • Skupianie (ang. clustering) – (CD4) • Oligomeryzacja (CD95, Fas) – tylko trimer oddziałuje z trimerycznymFasL. • Dopasowanie struktury białka do błony Greaves J. Mol. Membrane Biol, 2009; 26(12): 6779
Przykład: Regulacja wzrostu aksonu: • FGF2 – zasadowy czynnik wzrostu fibroblastów po związaniu ze swoim receptorem aktywuje jeden z enzymów PAT • Zachodzi palmitylacja dwóch izomerów neuronalnego białka adhezyjnego NCAM140 i NCAM180 co powoduje: • ich translokację do tratw lipidowych • wzrost własności adhezyjnych • wzrost aksonu
Prenylacja farnezylacja i geranylogeranylacja Enzymy prowadzące reakcje prenylacji białek: FT - farnezylotransferaza GGT I - geranylogeranylotransferaza I GGT II - geranylogeranylotransferaza II) = = geranylogeranylotransferaza Rab
miejsce działania statyn
Farnezylacji przez FT ulegają białka z sekwencją C-końca: -Cys-a-a-X-COO- a – aminokwas alifatyczny X – zwykle Met, Ser, Ala, Gln Geranylogeranylacji przez GGT I ulegają białka z sekwencją C-końca: -Cys-a-a-X-COO- X – zwykle Leu Geranylogeranylacji przez GGT II ulegają białka z sekwencjami C-końca: -Cys-Cys-COO- -Cys-X-Cys-COO-
Przebieg farnezylacji Geranylogeralynacja prowadzona przez GGT I zachodzi tak samo. Geranylogeranylacja przez GGT II sekwencja: CC – brak estryfikacji grupą metylową sekwencja CXC – zachodzi estryfikacja grupą metylową
Prenylacja a aktywność białek Brak prenylacji znacznie osłabia aktywność białek, które powinny ulegać tej modyfikacji. Ponieważ bardzo duża grupa nowotworów charakteryzuje się mutacją w genie Ras powodującą jego stałą aktywność, próbuje się stosować inhibitory prenylotransferaz w terapiach nowotworowych. Dobre wyniki in vitro, niezbyt zachęcające próby kliniczne. Statyny – inhibitory reduktazy HMG-CoA (3-hydroksy,3-metyloglutarylo-CoA HMG-CoA – enzym szlaku syntezy cholesterolu
Potranslacyjna modyfikacja N-końca Usunięcie metioniny • Zachodzi kotranslacyjnie • Reakcję prowadzi aminopeptydaza metioninowa • Preferowana jest hydroliza wiązania pomiędzy resztą Met a małym aminokwasem: G, A, S, C, T, P, V • Prolina w pozycji 3 może hamować tę reakcję • Usunięcie metioniny może warunkować aktywność biologiczną niektórych białek • Usunięcie metioniny może warunkować inną potranslacyjną modyfikację - mirystylację
ACETYLACJA I METYLACJA (wewnątrz cząsteczki białka) ACETYLACJA • Proces odwracalny – białka mogą ulegać acetylacji przez acetylotransferazy(HAC) i deacetylacji przez deacetylazy (HDAC) • Większość poznanych procesów acetylacji zachodzi w jądrze komórkowym • Acetylacji ulegają nieliczne reszty lizyny (sekwencja zgodności sprzyjająca acetylacji – niezdefiniowana) • Aktywność acetylaz jest regulowana przez sygnały prowadzące do proliferacji i różnicowania (np. poprzez fosforylację).
METYLACJA • Modyfikacja odwracalna prowadzona przez enzymy: białkowe metylotransferazy • Metylacji może ulegać C-końcowy aminokwas, czasem metylacja towarzyszy prenylacji (białka G (trimeryczne np. transducyna), Ras) lub reszta argininy lub lizyny • Metylacja może podlegać regulacji, metylacja może regulować aktywność białek • Istnieje metylotransferaza białkowa wchodząca w interakcję z receptorem (PRMT1 oddziałuje z receptorem IFN-a) • Pod wpływem NGF – wzrost stopnia metylacji białek komórkowych • Grupy białek ulegające metylacji: • podjednostka g trimerycznych białek G np. transducyny, • małe białka G: Ras, Rho • hnRNP • białkowa fosfataza PP2A • histony
Poliubikwitynacja Potranslacyjna modyfikacja prowadząca do degradacji białka Polega na kowalencyjnym połączeniu ubikwityny do grupy w NH2 lizyny białka. Do pierwszej przyłączonej Ub zostają przyłączone kolejne cząsteczki Ub. Łańcuch poliubikwitynowy przyłączony do białka jest znacznikiem dla degradacji przez proteasom. Ubikwityna 76 aminokwasów
Degradacja białek • Rozkład nienatywnych cząsteczek • Utrzymanie puli aminokwasów w sytuacji głodu • REGULACJA POZIOMU BIAŁEK Okres półtrwania dla niektórych białek to minuty, dla innych wiele dni. Białka krótkożyjące zawierają degrony – sygnały degradacji. Najlepiej poznanym degronem jest N-degron stanowiący po prostu N-końcowy aminokwas białka. Reguła N-końca (ang. the N-end rule): Jeśli N-końcowym aminokwasem białka jest: aminokwas zasadowy (Arg, Lys, His) lub duży aminokwas hydrofobowy (Leu, Ile, Trp, Tyr, Phe) to takie białko ulega szybkiej degradacji.
System Ubikwityna - Proteasom E1 – enzym aktywujący Ub E2 – enzymy przenoszące Ub E3 – ligazy białko:Ub
Ubikwitynacja nie zawsze prowadzi do degradacji Ubikwitynacja białek może wpływać na wiele procesów i funkcji białek
Sumoilacja SUMO – ang. small ubiquitin-like modifier 110 aminokwasów, niewielkie podobieństwo sekwencji aminokwasowej (18% identycznych aminokwasów), ale duże podobieństwo struktury z Ub.
Sumoilacji ulegają także reszty lizyny białek i proces przebiega podobnie do procesu ubikwitynacji. • Udział w podziałach komórkowych, proces istotny dla organizacji chromosomów mitotycznych. Mutacje letalne. • Regulacja ekspresji genów – w większości przypadków – zahamowanie: • - promuje oddziaływanie z ko-represorami - białkami modulującymi chromatynę • - wywołuje zmiany w konformacji i oddziaływaniu białek z DNA • - wpływa na lokalizację białek Sumoilacja czynników transkrypcyjnych może wzmagać (HSF1, p53) lub hamować ich aktywność (c-Jun, Elk-1).
Ubikwitynacja versus sumoilacja To, że w niektórych przypadkach ta sama reszta Lys ulega ubikwitynacji i sumoilacji pozwoliło na postawienie hipotezy, że w te dwa procesy mogą mieć przeciwne efekty. Ubikwitynacja histonów – aktywacja transkrypcji; sumoilacja – zahamowanie transkrypcji. PCNA (antygen jądrowy proliferujących komórek) może ulegać i ubikwitynacji i sumoilacji. Przypuszcza się, że te modyfikacje decydują o tym, która polimeraza prowadzi replikację – polimeraza d czy któraś z polimeraz Y (translesion synthesis).
Prawidowe zwijanie białek jest wspomagane przez białka opiekuńcze • wiele białek opiekuńczych należy do białek szoku cieplnego (heat shock proteins). • Hsp zostały poznane jako te, których ekspresja znacząco wzrasta przy wzroście temperatury. • Nie wszystkie białka opiekuńcze są syntetyzowane w zwięszonej ilości w sytuacjach stresowych np. Hsp70 jest, Hsc70 – nie jest. • Białka opiekuńcze rozpoznają nieprawidłowo sfałdowane białka - rozpoznają wyeksponowane fragmenty hydrofobowe. W białkach prawidłowo sfałdowanych fragmenty hydrofobowe znajdują się „wewnątrz” cząsteczki białka.
·Zbudowane z dwóch pierścieni. Każdy pierścień składa się z 7 takich samych lub bardzo podobnych białek – razem tworzących „beczkę”
·GroEL może przyjmować dwa stany: 1.wiążący – „beczka” jest otwarta, wnętrze „beczki” hydrofobowe 2.stan aktywny – w którym możliwe jest zachodzenie fałdowania. Stan aktywny jest indukowany związaniem ATP oraz – w przypadku Prokaryota - co-chaperoniny zbudowanej z 7 podjednostek (10kDa) tworzących także pierścień (GroES).Dramatyczna zmiana konformacji GroEL powierzchnia staje się hydrofilowa faworyzując schowanie hydrofobowych części białka-substratu do wnętrza molekuły i faworyzując ekspozycje miejsc hydrofilowych. ·Dwa pierścienie GroEL kooperują ze sobą. Związanie ATP do jednego pierścienia umożliwia oddysocjowanie GroES oraz uwolnienie sfałdowanego (lub nie!) białka