Download
1 / 1

E N D
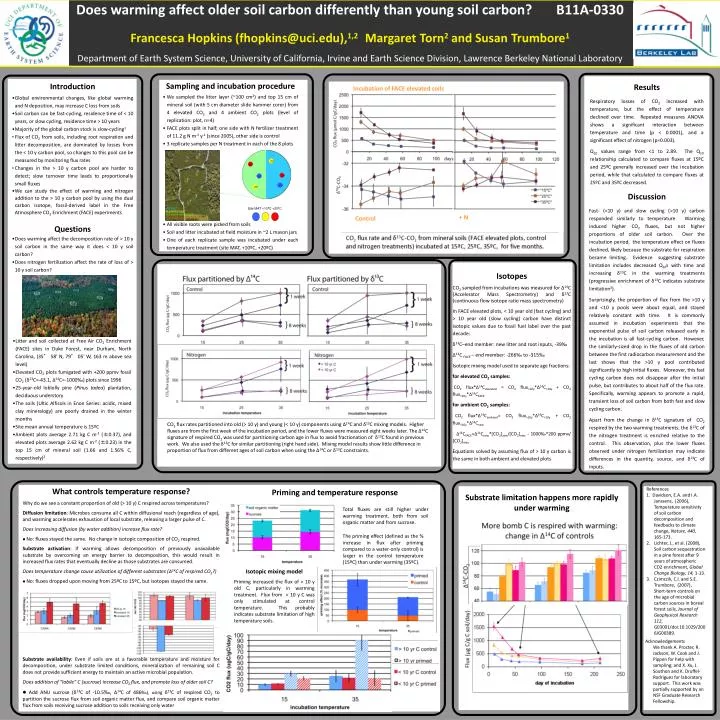
Discussion Fast- (< 10 y) and slow cycling (> 10 y) carbon responded similarly to temperature. Warming induced higher CO2 fluxes, but not necessarily higher proportions of older soil carbon. The temperature effect on fluxes declined, likely because the substrate for respiration became limiting. There are multiple lines of evidence to indicate substrate limitation, including decreased Q10s with time, and increasing δ13C in the two warming treatments over the course of the incubation support this (enrichment of δ13C during an incubation time course is often indicative of substrate limitation3). Mixing model calculation show little difference in the patterns and proportion of flux from different ages of soil carbon when using the Δ14C or δ13C constraints. Surprisingly, the proportion of flux from the > 10 y and < 10 y pools stayed relatively constant with time. It is commonly assumed in incubation experiments that the exponential pulse of soil carbon released early in the incubation is all fast-cycling carbon. However, the similarly-sized drop in the fluxes of old carbon between the first radiocarbon measurement and the last shows that the >10 y pool contributed to high initial fluxes. In particular, warming appears to promote a rapid, transient loss of soil carbon from both fast and slow cycling carbon. Apart from the change in δ13C signature of CO2 respired by the two warming treatments, the δ13C of the nitrogen treatment is enriched relative to the control. This observation, plus the lower fluxes observed under nitrogen fertilization perhaps indicate differences in the quantity, source, and δ13C of inputs. Does warming affect older soil carbon differently than young soil carbon? B11A-0330 Francesca Hopkins (fhopkins@uci.edu),1,2MargaretTorn2and SusanTrumbore1 Department of Earth System Science, University of California, Irvine and Earth Science Division, Lawrence Berkeley National Laboratory 25º 35º 35º 15º 15º 25º Site MAT +10ºC +20ºC • Introduction • Global environmental changes, like global warming and N deposition, may increase C loss from soils • Soil carbon can be fast-cycling, residence time of < 10 years, or slow cycling, residence time > 10 years • Majority of the global carbon stock is slow-cycling1 • Flux of CO2 from soils, including root respiration and litter decomposition, are dominated by losses from the < 10 y carbon pool, so changes to this pool can be measured by monitoring flux rates • Changes in the > 10 y carbon pool are harder to detect; slow turnover time leads to proportionally small fluxes • We can study the effect of warming and nitrogen addition to the > 10 y carbon pool by using the dual carbon isotope, fossil-derived label in the Free Atmosphere CO2 Enrichment (FACE) experiments • Questions • Does warming affect the decomposition rate of > 10 y soil carbon in the same way it does < 10 y soil carbon? • Does nitrogen fertilization affect the rate of loss of > 10 y soil carbon? c • Sampling and incubation procedure • We sampled the litter layer (~100 cm2) and top 15 cm of mineral soil (with 5 cm diameter slide hammer corer) from 4 elevated CO2 and 4 ambient CO2 plots (level of replication: plot, n=4) • FACE plots split in half; one side with N fertilizer treatment of 11.2 g N m-2 y-1 (since 2005), other side is control • 3 replicate samples per N treatment in each of the 8 plots • All visible roots were picked from soils • Soil and litter incubated at field moisture in ~2 L mason jars • One of each replicate sample was incubated under each temperature treatment (site MAT, +10ºC, +20ºC) Results Respiratory losses of CO2 increased with temperature, but the effect of temperature declined over time. Repeated measures ANOVA shows a significant interaction between temperature and time (p < 0.0001), and a significant effect of nitrogen (p=0.003). Q10 values range from <1 to 2.89. The Q10 relationship calculated to compare fluxes at 15ºC and 25ºC generally increased over the incubation period, while that calculated to compare fluxes at 25ºC and 35ºC decreased. Incubation of FACE elevated soils Nitrogen Control CO2 flux rate and δ13C-CO2 from mineral soils (FACE elevated plots, control and nitrogen treatments) incubated at 15ºC, 25ºC, 35ºC, for five months. Discussion Fast- (<10 y) and slow cycling (>10 y) carbon responded similarly to temperature. Warming induced higher CO2 fluxes, but not higher proportions of older soil carbon. Over the incubation period, the temperature effect on fluxes declined, likely because the substrate for respiration became limiting. Evidence suggesting substrate limitation includes decreased Q10s with time and increasing δ13C in the warming treatments (progressive enrichment of δ13C indicates substrate limitation3). Surprisingly, the proportion of flux from the >10 y and <10 y pools were about equal, and stayed relatively constant with time. It is commonly assumed in incubation experiments that the exponential pulse of soil carbon released early in the incubation is all fast-cycling carbon. However, the similarly-sized drop in the fluxes of old carbon between the first radiocarbon measurement and the last shows that the >10 y pool contributed significantly to high initial fluxes. Moreover, this fast cycling carbon does not disappear after the initial pulse, but contributes to about half of the flux rate. Specifically, warming appears to promote a rapid, transient loss of soil carbon from both fast and slow cycling carbon. Apart from the change in δ13C signature of CO2 respired by the two warming treatments, the δ13C of the nitrogen treatment is enriched relative to the control. This observation, plus the lower fluxes observed under nitrogen fertilization may indicate differences in the quantity, source, and δ13C of inputs. + N Control Isotopes CO2 sampled from incubations was measured for Δ14C (Accelerator Mass Spectrometry) and δ13C (continuous flow isotope ratio mass spectrometry) In FACE elevated plots, < 10 year old (fast cycling) and > 10 year old (slow cycling) carbon have distinct isotopic values due to fossil fuel label over the past decade: δ13C–end member: new litter and root inputs, -39‰ Δ14C FACE – end member: -266‰ to -315‰ Isotopic mixing model used to separate age fractions: for elevated CO2 samples: CO2 flux*Δ14Celevated = CO2 flux>10y*Δ14C>10y + CO2 flux<10y*Δ14CFACE for ambient CO2 samples: CO2 flux*Δ14Cambient= CO2 flux>10y*Δ14C>10y + CO2 flux<10y*Δ14Cnew Δ14CFACE=Δ14Cnew*[CO2]atm/[CO2]elev - 1000‰*200 ppmv/ [CO2]elev Equations solved by assuming flux of > 10 y carbon is the same in both ambient and elevated plots Site Description • Litter and soil collected at Free Air CO2 Enrichment (FACE) sites in Duke Forest, near Durham, North Carolina, (35° 58' N, 79° 05' W, 163 m above sea level) • Elevated CO2 plots fumigated with +200 ppmv fossil CO2 (δ13C=-43.1, Δ14C=-1000‰) plots since 1996 • 25-year-old loblolly pine (Pinus tadea) plantation, deciduous understory • The soils (Ultic Alfisols in Enon Series: acidic, mixed clay mineralogy) are poorly drained in the winter months • Site mean annual temperature is 15ºC • Ambient plots average 2.71 kg C m-2 (±0.37), and elevated plots average 2.62 kg C m-2 (±0.23) in the top 15 cm of mineral soil (1.66 and 1.56% C, respectively)2 CO2 flux rates partitioned into old (> 10 y) and young (< 10 y) components using Δ14C and δ13C mixing models. Higher fluxes are from the first week of the incubation period, and the lower fluxes were measured eight weeks later. The Δ14C signature of respired CO2 was used for partitioning carbon age in flux to avoid fractionation of δ13C found in previous work. We also used the δ13C for similar partitioning (right hand side). Mixing model results show little difference in proportion of flux from different ages of soil carbon when using the Δ14C or δ13C constraints. What controls temperature response? Why do we see a constant proportion of old (> 10 y) C respired across temperatures? Diffusion limitation: Microbes consume all C within diffusional reach (regardless of age), and warming accelerates exhaustion of local substrate, releasing a larger pulse of C. Does increasing diffusion (by water addition) increase flux rate? No: fluxes stayed the same. No change in isotopic composition of CO2 respired. Substrate activation: If warming allows decomposition of previously unavailable substrate by overcoming an energy barrier to decomposition, this would result in increased flux rates that eventually decline as those substrates are consumed. Does temperature change cause utilization of different substrates (Δ14C of respired CO2?) No: fluxes dropped upon moving from 25ºC to 15ºC, but isotopes stayed the same. Substrate availability: Even if soils are at a favorable temperature and moisture for decomposition, under substrate limited conditions, mineralization of remaining soil C does not provide sufficient energy to maintain an active microbial population. Does addition of “labile” C (sucrose) increase CO2 flux, and promote loss of older soil C? Add ANU sucrose (δ13C of -10.5‰, Δ14C of 486‰), using δ13C of respired CO2 to partition the sucrose flux from soil organic matter flux, and compare soil organic matter flux from soils receiving sucrose addition to soils receiving only water Priming and temperature response • References • 1. Davidson, E.A. and I.A. Janssens, (2006), Temperature sensitivity of soil carbon decomposition and feedbacks to climate change, Nature,440, 165-173. • Lichter, J., et al. (2008), Soil carbon sequestration in a pine forest after 9 years of atmospheric CO2 enrichment, Global Change Biology, 14, 1-13. • Czimczik, C.I. and S.E. Trumbore, (2007), Short-term controls on the age of microbial carbon sources in boreal forest soils, Journal of Geophysical Research112, G03001/doi:10.1029/2006JG00389. Substrate limitation happens more rapidly under warming Total fluxes are still higher under warming treatment, both from soil organic matter and from sucrose. The priming effect (defined as the % increase in flux after priming compared to a water-only control) is larger in the control temperature (15ºC) than under warming (35ºC). Isotopic mixing model Priming increased the flux of > 10 y old C, particularly in warming treatment. Flux from < 10 y C was only stimulated at control temperature. This probably indicates substrate limitation of high temperature soils. Acknowledgements We thank A. Procter, R. Jackson, W. Cook and J. Pippen for help with sampling; and X. Xu, J. Southon and K. Druffel-Rodriguez for laboratory support. This work was partially supported by an NSF Graduate Research Fellowship.