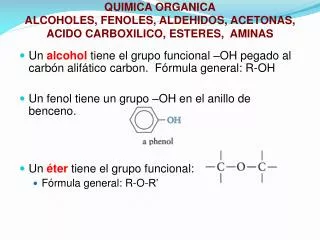
Download
1 / 32
460 likes | 1.92k Views
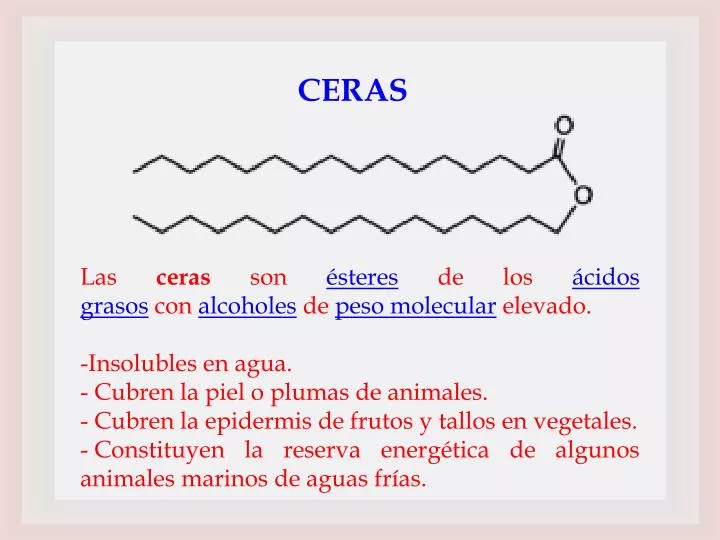
CERAS. Las ceras son ésteres de los ácidos grasos con alcoholes de peso molecular elevado. Insolubles en agua. Cubren la piel o plumas de animales. Cubren la epidermis de frutos y tallos en vegetales. Constituyen la reserva energética de algunos animales marinos de aguas frías.

E N D
CERAS • Las ceras son ésteres de los ácidos grasos con alcoholes de peso molecular elevado. • Insolubles en agua. • Cubren la piel o plumas de animales. • Cubren la epidermis de frutos y tallos en vegetales. • Constituyen la reserva energética de algunos animales marinos de aguas frías.
QUIMICA BIOLOGICA Lic. y Prof. en Cs. Biológicas BOLILLA 6: Lípidos. Digestión y absorción. Metabolismo de lípidos. Beta-oxidación. Ácidos grasos saturados, no saturados e insaturados de número par de átomos de C. Regulación en la utilización de sustrato. Oxidación de ácidos grasos de número impar de átomos de carbono. Oxidación peroxisómica de ácidos grasos. Rendimiento energético.Ciclo del Glioxilato. Localización. Importancia. Cuerpos cetónicos.
Degradación de Ácidos Grasos • Los AG cubren hasta el 40% de las necesidades totales de combustible en una dieta normal. • En períodos de ayuno prolongado, durante el ejercicio aeróbico sostenido, o durante la hibernación en ciertos animales, los AG son la única fuente de energía.
Hipoglucemia y/o estrés Adrenalina Glucagón Lipasa Hormona Sensible
Degradación (β-Oxidación) de Ácidos Grasos • Ocurre en tejidos como: Hígado, músculo esquelético, corazón, riñón, tejido adiposo, etc. • Comprende la oxidación del carbono βdel ácido graso. • Se lleva a cabo en las MITOCONDRIAS. • Requiere: • Activación del ácido graso en el citosol (requiere energía en forma de ATP) • Transporte al interior de la mitocondria
1) Activación del ácido graso • Ocurre en el Citosol. • La reacción es catalizada por la TIOQUINASA o Acil CoA sintetasa. • El pirofosfato es hidrolizado por una PIROFOSFATASA (esto hace que la reacción sea irreversible).
Acil-CoA Acil-CoA Acil-Carnitina -oxidación
Mitocondria Acil-CoA del paso de activación Acil-CoA deshidrogenasa Acetil-CoA Cetoacil-CoA tiolasa Enoil-CoA hidratasa 3-HO-Acil-CoA deshidrogenasa
β- Oxidación de Ac. Grasos • Los AG de cadena larga son procesados por las mismas 4 etapas cíclicas. • Por ciclo, se eliminan por oxidación sucesiva, 2 carbonos a partir del extremo carboxílico. • Se produce una molécula de Acetil-CoA en cada ciclo. • El acetil-CoA producido entra en el ciclo de Krebs para producir energía, oxidándose a CO2 y H2O.
En cada ciclo se pierden 2 átomos de C en forma de Acetil-CoA. • Para degradar completamente un AG de 16 C hacen falta : • 7 ciclos de β-Oxidación. • Nº de ciclos = (nº de C) – 1 • 2 • En cada ciclo se produce 1 molécula de FADH2 y otra de NADH +H: • FADH2= 2 ATP • NADH+H= 3 ATP
INTERRELACION CON EL CICLO DE KREBS • Los acetilos formados en la b-OXIDACIÓN ingresan al CICLO DE KREBS para su oxidación total a CO2. • Los NADH y FADH2 producidos en el CICLO DE KREBS forman ATP en la mitocondria (FOSFORILACIÓN OXIDATIVA)
BALANCE ENERGÉTICO PRODUCCIÓN DE ATP EN LA BETA- OXIDACIÓN 7 MOLÉCULAS DE FADH2 2 ATP X 7 = 14 ATP 7 MOLÉCULAS DE NADH 3 ATP X 7 = 21 ATP SUBTOTAL 35 ATP 8 MOLECULAS ACETIL CoA 12ATP X 8 = 96 ATP SUBTOTAL = 131 ATP ACTIVACIÓN DE PALMITATO A PALMITOIL CoA = - 2 ATP TOTAL 129 ATP
Oxidación de ácidos grasos de número impar de átomos de C Los ácidos grasos con número impar de átomos de C son oxidados por la vía de la β-oxidación produciendo restos de Acetil-CoA, hasta que en el último ciclo, el resto de 5 carbonos que se oxida, da lugar a Acetil-CoA (2C) más Propionil-CoA (3C). Ciclo de Krebs
Oxidación peroxisómica de ácidos grasos de cadena muy larga (C20 y C22) Energía disipada como calor Mitocondria Un nivel elevado de grasas en la dieta produce aumento de la actividad oxidativa en el peroxisoma. Esta organela no tiene las enzimas para degradar acetil CoA a CO2 . Octanoil-CoA + Acetil-CoA en última vuelta
Regulación de la utilización de sustrato por oxidación de ácidos grasos • Diferentes mecanismos reguladores evitan el gasto excesivo de sustratos y energía. • En el caso de la oxidación de los ácidos grasos, la célula posee mecanismos que permiten activar ó inhibir distintas vías relacionadas a fin de proveer las necesidades de la célula, según los diferentes estados metabólicos de la misma. • Por ejemplo, cuando la provisión de ácidos grasos en músculo es abundante, la utilización y oxidación de la glucosa se deprime. deshidrogenasa
Regulación de utilización de sustrato en la oxidación de ácidos grasos La entrada de los AG a la mitocondria es la etapa limitante de la oxidación
Durante la combustión completa de un AG se produce gran cantidad de agua, que deriva de las reacciones de fosforilación asociadas. Así, los camellos obtienen agua por degradación de sus reservas grasas El oso pardo entra en periodo de letargo por períodos de hasta 7 meses. Mantiene su temperatura corporal a31-33 C unos 5 grados menos que la normal. Consume 6000Kcal/día durante dicho periodo
Ciclo del glioxilato ¿Es posible sintetizar GLU a partir de Acetil-CoA? Las células animales NO son capaces de realizar la síntesis neta de azúcares a partir de Acetil-CoA. - Solo las plantas y algunas bacterias-
CICLO DEL GLIOXILATO Plantas, invertebrados y algunos microorganismos. En los vegetales, el ciclo tiene lugar en los Glioxisomas, organelas especializadas en las cuales se lleva a cabo la degradación de los ácidos grasos (β-oxidación) para producir Acetil-CoA que será utilizada en el ciclo. El Ciclo del Glioxilato comparte algunas enzimas del Ciclo de Krebs, pero incluye dos enzimas específicas localizadas en los glioxisomas. En cada vuelta del ciclo se utilizan 2 moléculas de Acetil-CoA para generar una de oxalacetato.
B-oxidacion de acidos grasos Sacarosa Glucosa Glioxisomas Ciclo del Glioxilato Glioxisoma Succinato Mitocondria
CICLO DEL GLIOXILATO Se evitan las 2 reacciones de descarboxilación del Ciclo de Krebs. El succinato formado en la reacción de la isocitrato liasa se transporta desde el glioxisoma a la mitocondria. Allí se convierte en oxalacetato por las reacciones del Ciclo de Krebs. De esa forma se puede utilizar para la síntesis de hidratos de carbono a través de la gluconeogénesis. • En el caso de las plantas, cuando las semillas germinan, los triglicéridos se degradan a glicerol (precursor de gluconeogenesis) y ácidos grasos que se degradan a Acetil-CoA (precursor del ciclo del glioxilato) y se convierten en azúcares, que aportan Energía para el crecimiento del vegetal.
Formación de Cuerpos Cetónicos (Cetogénesis) • Después de la degradación de los AG, Acetil-CoA es oxidado en el Ciclo de Krebs. • Para esto es necesaria la presencia de oxalacetato (1er intermediario del ciclo de Krebs). • Si la cantidad de este es insuficiente, las unidades de acetil-CoA son utilizadas mediante una vía alternativa en la que se producen los Cuerpos Cetónicos: Acetona, Acetoacetato y 3-Hidroxi-butirato.
CUERPOS CETÓNICOS Son compuestos derivados de acetil CoA que, al acumularse y no entrar al ciclo de Krebs es convertidos en cuerpos cetónicos. El hígado es el principal productor ya que posee todas las enzimas necesarias. Es incapaz de usarlos como combustible. Los órganos que los usan son: cerebro, músculo esquelético, corazón y otros tejidos que no los sintetizan. Solo se usan como fuente de energía en situaciones metabólicas especiales. Ej.: Diabetes, ayuno prolongado, dieta grasa. El aumento de estos provoca Acidosis Metabólica. La cetogénesis se produce en las mitocondrias hepáticas.
CETOGENESIS Precursor • El 1er paso es la inversa de la última etapa de la b-oxidación. • Acetoacetil- CoA se condensa con otro acetil-CoA para dar HMG-CoA. • El HMG-CoA se rompe formando acetoacetato y Acetil-CoA. • El Acetoacetato puede originar los otros cuerpos cetónicos.
Formación y exportación de cuerpos cetónicos Gotas de lípidos • Los cuerpos cetónicos se forman y exportan desde el hígado. • En condiciones energéticamente desfavorables (diabetes, inanición), oxalacetato deriva hacia la gluconeogénesis, para liberar glucosa a la sangre. • El ciclo de Krebs trabaja muy lentamente. Hepatocito Acetoacetato y b-hidroxi- butirato exportados como energía para: corazón, músculo, riñón y cerebro. Glucosa exportada como combustible para cerebro y otros tejidos.
UTILIZACIÓN DE LOS CUERPOS CETÓNICOS Ocurre en tejidos EXTRAHEPÁTICOS • Los tejidos extrahepáticos utilizan cuerpos cetónicos como fuente de energía. • Acetil CoA dentro de la célula ingresa al ciclo de Krebs para obtener energía. Ausente en hígado
Bibliografia 1- BLANCO A., “Química Biológica”, Ed. El Ateneo, 8a edic., Bs. As. (2007). 2- LEHNINGER, A.L., "Principios de Bioquímica", Ed. Omega, 4ª ed. (2008). 3- Docentes de Química Biológica, “QUIMICA BIOLOGICA Orientada a Ciencias de los Alimentos”, Nueva Editorial Universitaria de la Universidad Nacional de San Luis. 4- MURRAY R y col., “Bioquimica de Harper”, Ed. El Manual Moderno, 14º ed. (1997). Bibliografía Complementaria 1- CAMPBELL Y FARREL, “Bioquimica”, Thomson Eds., 4ta. Ed., (2005). 2- LIM M.Y., “ Lo esencial en Metabolismo y Nutrición”, Ed. Elsevier, 3ra. ed., Barcelona (2010).