Download

1 / 32
490 likes | 949 Views
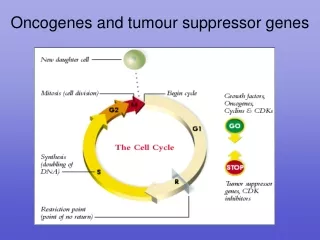
Tumour Suppressor Genes. TSGs code for proteins that inhibit cell division Mutations can cause the proteins to be inactivated and may thus deprive cells of needed restraints on proliferation. Ways of inactivating a TSG. Active growth promoter. S. P53 – protein of 53kDa Rb

E N D
Tumour Suppressor Genes TSGs code for proteins that inhibit cell division Mutations can cause the proteins to be inactivated and may thus deprive cells of needed restraints on proliferation
Ways of inactivating a TSG Active growth promoter S P53 – protein of 53kDa Rb - retinoblastoma Inactive growth complex S Active growth promoter Active growth promoter Active growth promoter Mdm2 P S S S Binding protein phosphorylation mutation
p53 – ‘guardian’ of the genome • Initially identified as a tumour specific nuclear antigen of 53kDa • 4 monomers form a functional tetramer • Comparison with normal cells showed the presence of mutations in cancer • When wild-type gene transfected into tumours, it stopped their growth i.e. a tumour suppressor gene Cellular stress Cell proliferation p53 p53 Stimulates DNA repair p53 p53 Prepares for apoptosis
p53 mutations and cancer • 50% of all cancers show mutations in p53 • 90% mutations in Squamous Cell Carcinoma (SCC) • 80% point mutations and 20% truncations • Mutations in p53 • cause loss of function • Leads to continued cell division despite DNA damage • Leads to increased mutation rate Axillary lymph node infiltrated by metastatic breast carcinoma: IHC for p53 protein
P53 – domain structure Transactivation domain Tetramer formation C-terminal domain DNA binding 100 200 300 393 1
P53 – posttranslational modification sites Transactivation domain Tetramer formation DNA binding C-terminal domain circles, Ser/Thr phosphorylation sites; hexagons, acetylation sites; octagons, sumoylation site. post-translational modifications are induced when cells are exposed to stress The N-terminus is heavily phosphorylated while the C-terminus contains phosphorylated, acetylated and sumoylated residues. N-terminal phosphorylations are important for stabilizing p53 and are crucial for acetylation of C-terminal sites, which in combination lead to the full p53-mediated response to genotoxic stresses.
P53 – Transcriptional activation Transactivation domain Tetramer formation C-terminal domain DNA binding stimulates transcription indirectly by binding to other nuclear proteins e.g.Mdm2, GADD45, Cyclin G, BAX, IGF –BP3
Mdm2 regulates p53 murine double minute 2 (mdm2) gene was originally identified by amplification in a spontaneously transformed mouse BALB/c cell line The Mdm2 protein binds to p53 and inhibits p53-mediated transcription activation Gene amplification of the human mdm2 gene, hdm2, observed in about a third of human sarcomas that retained wild-type p53 Overexpression of hdm2 is an alternative way of inactivating p53
Regulation of p53 by Mdm2 • Mdm2 inhibits the transcriptional activity of p53 by binding to the TAD. • 2) p53 is degraded by Mdm2. Mdm2 ubiquitinates p53 and itself, leading to the degradation of both proteins
genotoxic stress stabilises p53 After stress, p53 accumulates by the dissociation of the p53–Mdm2 interaction due to either 1) Posttranslational modification of p53and/or mdm2 (DNA damage response) 2) Other interacting proteins like ARF (alternative reading frame; p14ARF in human, p19ARF in mice). ARF sequesters Mdm2 to the nucleoli, stabilising the p53 tetramer (oncogenic stimulus and viral transformation) Oncogenic activation DNA damage Mdm2 P ARF p53 p53 p53 53
e.g. de novo glioblastoma multiforme (a.k.a grade IV astrocytomas) grow rapidly, invade nearby tissue and contain cells that are very malignant. clear MRI – death in < 1 year Caused by amplification of Mdm2 leading to p53 inactivation mdm2 and cancer In many cancers, the protective activity of p53 is switched off, as a result of overexpression of the protein MDM2, which binds to the transactivation domain of p53 and blocks its ability to activate transcription
Dynamics of the p53-Mdm2 feedback loop in individual cells • p53-CFP (green) in clonal MCF7 cells after 5-Gy-irradiation. Time (min) post radiation shown • (d) Dynamics of p53-CFP (green) and Mdm2-YFP (red) in a cell that shows two pulses. Nature Genetics36, 147 - 150 (2004)
P53 – DNA binding domain Transactivation domain Tetramer formation Transcriptional inhibition DNA binding Sequence specific DNA binding: P53 acts as a transcription factor by binding to genes that have a p53 response element
Normal cells, p21 is not transcribed Upon DNA damage, p53 levels are increased, which in turn actively transcribe the p21 gene p21 represses cell growth by binding to Cdk's (cyclin-dependent protein kinases) in the cell resulting in G1 arrest Mutations in the DNA binding domain of p53 results in loss of DNA binding affinity. Subsequently, p53 can no longer induce transcription of p21. If there is no p21, there is no way to bind the Cdk's and the cell proceeds, uncontrolled, through the cell cycle. P53 – DNA binding
P53 – tetramer formation Transactivation domain Tetramer formation C-terminal domain DNA binding
P53 – tetramer formation Children in southern Brazil exhibit an elevated incidence of adrenocortical carcinoma (ACC) due to an arginine to His mutation at 337 (R337H) within tetramerization domain of p53 (35/36 patients). Nature Structural Biology9, 12 - 16 (2001)
P53 – C-terminal domain Transactivation domain Tetramer formation C-terminal domain DNA binding The C-terminal regulatory domain has 2 functions Negative regulation: Phosphorylation destabilises the folding of the DNA binding domain Positive regulation:Acetylation of the C-terminus of DNA-bound p53 stabilises p300 binding, which is required for p53 driven transcription • It also modulates • the stability, • the oligomerization state, • the nuclear import/export process and • the degree of ubiquitination of p53.
p53 and ageing P53-/- mice develop normally but prone to cancers Overexpressing p53 mice are resistant to tumours but show premature ageing Nature 415, 26-27 (3 January 2002)
Retinoblastoma Rare childhood cancer of the eye (retinomas) that develops in children, typically under five years old. Incidence • 2 % of childhood malignancies Influencing factors 30-40% hereditary 60-70% sporadic Treatment Surgery, radiation, chemotherapy leukocoria, a white reflex of the pupil
Molecular genetics of Rb The 2-hit hypothesis Nondisjunction & duplication Mitotic recombination deletion Point mutation
Retinoblastoma protein (pRb) • Normally inhibits cell proliferation • localised in the nucleus • tumour suppressor protein of ~110kD • pRb has > 10 phosphorylation sites (affects protein-protein interaction) • Rb gene is 300kb long & mutations in this gene leads to loss of function. • Most mutations involve gross chromosomal changes in the 3kb coding region and 1/3 are point mutations. • Loss of heterozygosity at chromosome 13q14.2.
Endogenous expression and phosphorylation of Rb in human neuronal cultures
Other TSGs Genes for cytoplasmic proteins APC Involved in colon & stomach cancers DPC4 Involved in pancreatic cancers. Codes for signalling molecule involved in inhibition of cell division NF-1 Codes for inhibitor of ras protein. Involved in neurofibroma, pheochromocytoma (peripheral nervous system) & myeloid leukaemia NF-2 Involved in meningioma & ependynoma (brain) & schwannoma (shwann cells surrounding the neuron) Genes for nuclear proteins (also includes Rb and p53) MTS1 Codes for the p16 protein, a CKI involved in a range of cancers. WT1 Codes for transcription factor WT1, involved in Wilms' tumor Genes with unclear cellular locations BRCA1/2 Involved in breast/ovarian cancers (DNA repair?) VHL Involved in renal cell cancer
Some questions • Why do you get mainly retinal cancer when the Rb gene is knocked out? • Is the balance between activity of oncogenes and tumour suppressor genes a cumulative effect? • Is cancer a disease not of cell division but of cell differentiation? NATURE|VOL 427 | 15 JANUARY 2004 |www.nature.com/nature
References • Chapter 14 Cell & Mol Biol by Knowles and Selby AND/OR • Chapter by RJB King