Download

1 / 15
150 likes | 326 Views
DNA Structure and Replication. DNA is the Genetic Material.

E N D
DNA is the Genetic Material • DNA and RNA were first described by Friedrich Miescher in 1869. He isolated a phosphorus-containing material from the nuclei of cells found in pus from discarded surgical bandages, and called in “nuclein”. He later found the same material in salmon sperm. Later it was recognized that DNA and RNA are slightly different in structure. • In pre-digital days it seemed impossible for complex phenotypic traits to be coded in a simple linear fashion. DNA was known to be a linear polymer of just 4nucleotides, and it was thought to be just a scaffold for the actual genes. Proteins were considered the most likely genetic material, or perhaps some undiscovered substance. • Definitive proof of DNA’s central role came in the 1950’s, but important experiments were done earlier.
Transformation Experiments • In 1928, F. Griffith did a series of experiments on mice infected with Streptococcus bacteria. These experiments were followed up Avery, MacLeod, and McCarty in 1943, who demonstrated that at least in this case, DNA was the hereditary material, and not proteins. • Griffith had 2 strains of Streptococcus. The S strain had a polysaccharide coat around each cell, causing the colonies to have a smooth, glossy appearance. The R strain lacked the coat, and its colonies had a rough appearance. More importantly, the S strain was virulent: when injected into mice, they developed pneumonia and died. The R strain was avirulent: it did not kill the mice upon injection. • When S cells were killed by heat, injecting them had no effect on the mice. Heat killed R cells also had no effect. • The surprising result: when live R cells were mixed with heat-killed S cells and injected, the mice developed an infection and died. When bacteria were isolated from the dead mice, they were found to be S type. • Conclusion: something from the dead S cells had “transformed” the live R cells into S.
More Transformation • Avery, MacLeod, and MacCarty carried this result further by fractionating the dead S cells. They mixed various components (such as lipids, polysaccharides, protein, nucleic acids) of the S cells with live R cells, to determine which component caused the transformation. They found the DNA by itself caused the transformation, and no other component had any effect. • This demonstrated that DNA was the hereditary material, but their results were not considered to be generally applicable to inheritance.
Hershey-Chase Experiment • Another experiment, performed by Hershey and Chase in 1952, demonstrated the essential role of DNA using bacteriophage. At the time, a group of physicists who had previously worked on nuclear energy were moving into biology. They started work on the genetics of bacteria and bacteriophage that grew into modern molecular biology. • An important element of the Hershey-Chase experiment is that DNA contains phosphorus but not sulfur, while protein contains sulfur but not phosphorus. Thus it is possible to label the two types of molecule independently, with radioactive 32P and 35S. • Phage contain only DNA and protein, and no other types of molecule. • They used phage T2, infecting E. coli. • Hershey and Chase showed that 32P-labelled DNA entered the bacterial cells when the phage infected them, and that the new generation of phage contained a significant amount of that labelled DNA. • In contrast, the 35S-labelled protein stayed outside the cells during an infection, and none of it ended up in the new phage. • This implies that DNA is necessary for phage replication.
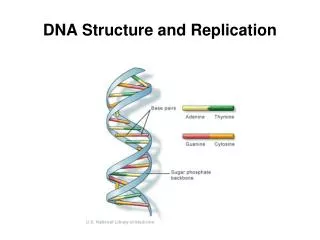
Structure of DNA • Once the importance of DNA was recognized, it was necessary to deduce how the DNA molecule is structured. A race between various lab groups ensued, and in 1953 James Watson and Francis Crick published a model of DNA structure. Their work was based on X-ray crystallography data provided by Maurice Wilkins and Rosalind Franklin. • DNA consists of two anti-parallel chains twisted into a helix. The nitrogenous bases are paired in the center of the molecule, and the phosphate-sugar backbones are on the outside. • Although DNA is the genetic material of all living cells, some viruses use RNA as their genetic material.
Nucleotide Structure • DNA and RNA are macromolecules composed of subunits called nucleotides. • Each nucleotide of DNA or RNA has 3 parts: a nitrogenous base, a sugar, and a phosphate group. • The phosphate group, PO4, links two sugar molecules in the backbone. Each phosphate carries a -1 charge. This causes DNA to have an overall negative charge. • The sugar is ribose in the case of RNA and deoxyribose in the case of DNA. has 5 carbons, numbered 1’ through 5’. • the nitrogenous base is attached to the 1’ carbon • the 2’ carbon has a free -OH group in the case of RNA, but a -H group in the case of DNA. The lack of the oxygen atom makes DNA far less reactive than RNA. • the 3’ carbon has an -OH group on it that links to the phosphate group on the next base. The “end” of the DNA molecule is a free 3’ OH group. • the 5’ carbon is attached to the phosphate group.
More Nucleotide Structure • There are 4 possible DNA bases: adenine (A), guanine (G), cytosine (C), and thymine (T). • Adenine and guanine are purines: they consist of two linked rings of mixed nitrogen and carbon atoms. • Thymine and cytosine are pyrimidines, which consist of a single ring. In RNA, thymine is replaced by uracil (U), which looks like thymine except for a single methyl group. • Each strand of DNA pairs with a complementary DNA strand. This pairing happens because each A is paired with a T, and each G is paired with a C. Thus, the information on one DNA strand easily allows the other strand to be deduced. The amount of A in DNA always equals the amount of T, and the amount of G always equals the amount of C. This is not true in RNA, which is usually single-stranded. • Pairing is caused by hydrogen bonds, weak links between oxygen and nitrogen atoms where one of them has a hydrogen attached. • A-T pairs have 2 hydrogen bonds, while G-C pairs have 3 hydrogen bonds. G-C pairs are stronger, and they are more frequent in high temperature organisms.
Semi-conservative Replication • Watson and Crick recognized that the double stranded DNA molecule could replicate by unwinding, then synthesizing a new strand for each of the old stands. • This mode of replication is called “semi-conservative”. It means that after one DNA molecule has replicated to become 2 DNA molecules, each new molecule consists of one old strand (from the original molecule) and one new strand. • The information from each old strand can be used to create the new strands, since A always pairs with T, and G always pairs with C. • DNA replication starts at specific locations “origins of replication”, and proceeds in both directions.
Replication Components • The raw materials of DNA synthesis are “nucleoside triphosphates”, often written as “dNTPs”. • dNTPs have a chain of 3 phosphate groups attached to the 5’ carbon of the deoxyribose sugar. Just as with ATP, the bonds between the phosphates are high energy bonds, and releasing them produces the energy needed to drive the synthesis of DNA. • Each new nucleotide is added to a growing DNA chain by removing the outer 2 phosphates and attaching the remaining phosphate to the 3’ OH group of the previous nucleotide. • The DNA chain is said to grow from 5’ to 3’, which means that the first DNA base has a free 5’ end, with attached phosphates. The last nucleotide has a free 3’ OH group on it. All other bases have their 5’ carbons attached to a phosphate, which is attached to the 3’ OH group of the previous nucleotide. • DNA polymerase is the main enzyme used to replicate DNA. However, DNA polymerase is only one enzyme in the replication complex. Several other enzymes are needed to cause replication to occur.
Continuous and Discontinuous Synthesis • DNA can only be synthesized from 5’ to 3’, by adding new nucleotides to the 3’ end. • This is a problem, because both strands must be synthesized at the replication fork, and one strand will necessarily be synthesized in the opposite direction from the movement of the replication fork. • In reality, one strand is synthesized continuously, in the same direction that the replication form is moving. This is called the “leading strand”. • The other strand is synthesized in short, discontinuous pieces, that are then attached together to form the final DNA strand. This is the “lagging strand”. Each fragment of the lagging strand is called an “Okazaki fragment”, and they are synthesized in the opposite direction that the replication fork moves.
Discontinuous Synthesis • Another peculiarity of DNA synthesis is that DNA polymerase must attach new bases to the 3’ end of a pre-existing nucleic acid chain. All DNA synthesis starts at a short double-stranded region. • In the cell, short pieces of RNA, called “primers” are paired with the DNA bases to create to the short double stranded regions that DNA synthesis builds. • The RNA primers are synthesized by an enzyme called “primase”, and they are removed by DNA polymerase during the synthesis of the next Okazaki fragment. • Joining of the Okazaki fragments is done by the enzyme “DNA ligase”.
Replication Miscellany • Errors occur in DNA replication fairly frequently: the wrong base gets inserted due to the peculiarities of nucleotide chemistry. However, DNA polymerase has a built-in editing function that removes most of the incorrect bases. • Enzymes that replicate the RNA genomes of some viruses do not have editing functions. Thus, mutation rates in RNA viruses are 100-1000 times higher than in DNA viruses. This rapid mutation rate makes it easy for RNA viruses to evade the immune system. • DNA in cells is “supercoiled”, twisted into tight knots because the helix is given a few extra twists beyond what it needs to maintain its shape. Supercoiling has the advantage of causing the DNA to be more compact inside the cell, but it must be created and maintained by various enzymes that wind the DNA, break and rejoin DNA strands so they can pass through one another in the winding process, and stabilize single stranded molecules.