Download

1 / 103
1.05k likes | 1.51k Views
Sistema de Endomembranas. Biología Celular y Bioquímica I Esteban Osorio Cadavid Universidad del Valle. ENDOMEMBRANAS CELULARES ENDOMEMBRANAS . Las membranas internas o endomembranas de las células eucarióticas son las responsables de su compleja compartimentación.

E N D
Sistema de Endomembranas Biología Celular y Bioquímica I Esteban Osorio Cadavid Universidad del Valle
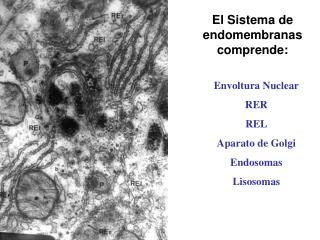
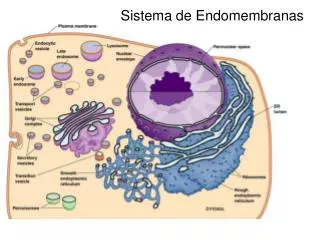
ENDOMEMBRANAS CELULARES ENDOMEMBRANAS. Las membranas internas o endomembranas de las células eucarióticas son las responsables de su compleja compartimentación. Las organelas son espacios con identidad bioquímica y funcional, delimitados por membranas lipoproteicas a través de las cuales deben pasar las sustancias que entran o salen de ellos. Entre varios de estos compartimentos se presentan un tráfico intenso de moléculas, realizado en muchas ocasiones por intermedio de vesículas. Obviamente, el desprendimiento y la fusión de estas vesículas tiene que ver con la estructura y con la composición de estas endomembranas. El retículo endoplásmico, el aparato de Golgi, los endosomas, los lisosomas y los microcuerpos son componentes esenciales del tráfico vesicular entre los diferentes compartimentos celulares.
El reticulo endoplasmático • Se divide en dos: • 1. El retículo endoplasmático rugoso, RER • 2. El Retículo endoplasmático liso, REL
RETÍCULO ENDOPLASMICO Se denomina así al conjunto de cisternas y canales tubulares citoplasmáticos descubierto por Porter, Claude y Fulman en 1945. Se reconocen dos tipos de retículo endoplasmático (RE), que se distinguen morfológica y funcionalmente: 1. Retículo endoplasmático rugoso (RER), con ribosomas adheridos a sus membranas y funcionalmente asociado a la ruta de secreción de proteínas. 2. Retículo endoplasmático liso (REL), sin ribosomas y asociado a la síntesis de lípidos y a la producción de membranas.
Los dos tipos de retículos coexisten en las células y a menudo sus cavidades están interconectadas. Las primeras aproximaciones a las funciones del RE vinieron de la observación de su abundancia relativa en determinadas células especializadas. El RER es muy abundante en las células que secretan proteínas, como por ejemplo las células pancreáticas, secretoras de enzimas digestivas y hormonas polipeptídicas. El REL se presenta muy desarrollado en células productoras de esteroides y otros tipos de lípidos. Por ejemplo, es muy abundante en las células testiculares intersticiales, que elaboran y secretan testosterona (una hormona esteroide).
Una segunda aproximación resulta de la fragmentación del RE durante los procedimientos de fraccionamiento celular. El RE queda reducido a numerosas vesículas de alrededor de 15 nm de diámetro, denominados MICROSOMAS. Los microsomas que provienen del RER presentan ribosomas adheridos a sus membranas. Los microsomas se han aislado para el estudio de su contenido enzimático y molecular. Se han detectado numerosas enzimas que han proporcionado evidencias importantes sobre las reacciones metabólicas que ocurren en el RE.
RETÍCULO ENDOPLASMICO RUGOSO (RER) Sus cisternas tienen de 20 a 100 nm de espesor y, como ya se indicó, sus membranas presentan una marcada asimetría: presentan ribosomas adheridos por el lado citoplasmático o citosólico. Los ribosomas se anclan a través de la subunidad grande a sitios definidos, distantes unos 30 nm uno del otro. Se ha identificado un receptor ribosomal en las membranas del RER de células de mamíferos, el cual consiste de una proteína transmembranal de 180 Kdal con un gran dominio citosólico de 160 Kdal, al que se asocia la subunidad grande de los ribosomas. En las membranas del RER también se han encontrado las proteínas denominadas RIBOFORINAS, de 63 a 65 Kdal, que parecen tener relación con el anclaje de los ribosomas.
Experimentos de “pulso y caza”, utilizando aminoácidos marcados radiactivamente, efectuados por Caro y Palade en células pancreáticas de conejillos de indias, mostraron la relación de RER con la síntesis, transporte y secreción de proteínas a través del Aparato de Golgi. Estos investigadores suministraron los aminoácidos radiactivos por 3 minutos (pulso). Al cabo de este tiempo, la radiactividad sólo fue detectada en las cavidades de RER (caza). A continuación, se suspendió la administración de los aminoácidos radiactivos, los cuales fueron sustituidos por aminoácidos no radiactivos. A los 17 minutos, la radiactividad se detectó en el RER próximo al aparato de Golgi y en las vesículas relacionadas con éste. A los 117 minutos, la marca radiactiva sólo era detectable en los “gránulos de zimógeno”, vesículas secretoras de enzimas provenientes del aparato de Golgi.
Sin embargo, no todas las proteínas celulares entran en las cavidades del RER, ni son llevadas al aparato de Golgi. Muchas son sintetizadas por los ribosomas en el citosol y se quedan allí o son transportadas al núcleo, cloroplastos, mitocondrias, microcuerpos o a otras organelas. Se ha demostrado que para que las proteínas sean introducidas a las cavidades del RER, deben poseer una secuencia específica en su extremo amino, denominada SECUENCIA SEÑAL o PEPTIDO SEÑAL. En la mayoría de las proteínas examinadas, el péptido señal consta de aproximadamente 30 aminoácidos, de los cuales 4 a 12 tienen cadenas R poco polares (Glicina, leucina, isoleucina, valina, triptófano o fenilalanina). La necesidad de las secuencias señal para la entrada de proteínas a diversos compartimientos celulares, se ha convertido en una de las teorías unificadoras más importantes de la Biología Celular y Molecular.
El péptido señal es reconocido por una partícula denominada PARTICULA DE RECONOCIMIENTO DE LA SEÑAL (PRS), la cual ha podido aislarse removiendo las proteínas asociadas a los microsomas provenientes del RER luego de la remoción de los ribosomas. La PRS es una asociación de 6 polipéptidos y un ARN específico de 300 nucleótidos de longitud (ARN 7S).
El papel de la PRS en la entrada de las proteínas a las cavidades del RER, se muestra a través de los siguientes esquemas experimentales: MICROSOMAS DEL RER ribosomas VESÍCULAS SIN RIBOSOMAS NaCl 0.5 M VESÍCULAS “DESNUDAS” SISTEMA DE SÍNTESIS DE PROTEÍNAS “LIBRE DE CÉLULAS” LAS PROTEÍNAS SE SINTETIZAN Y QUEDAN POR FUERA DE LAS VESICULAS Pero si al sistema de síntesis de proteínas se añaden las proteínas removidas con el NaCl 0.5 M, entre ellas la PRS, las proteínas sintetizadas, son incorporadas cotraduccionalmente (simultáneamente, a medida que se van sintetizando) a las vesículas.
Aproximaciones experimentales adicionales, han permitido diseñar una teoría coherente sobre la función de la PRS; particularmente después de que se logró el aislamiento del receptor de la PRS, una proteína de 650 aminoácidos, situada en las membranas del RER. Por ejemplo, utilizando RNA mensajero de la proteína prolactina en un sistema de síntesis de proteínas libres de células de germen de trigo, en la presencia y ausencia de microsomas, se han obtenido los siguientes resultados: 1. Sistema sin PRS, ni receptor de PRS, ni microsomas: Se sintetiza la prolactina completa, con su péptido señal 2. Sistema con PRS, pero sin receptor de PRS, ni microsomas: La síntesis de la prolactina se inicia, pero se detiene cuando el polipéptido alcanza entre 70 y 100 aminoácidos de longitud. 3. Sistema con PRS, con receptor de la PRS soluble, pero sin microsomas: La prolactina se sintetiza completamente, con su péptido señal. 4. Sistema con PRS y con microsomas que poseen el receptor de la PRS en sus membranas: La prolactina se sintetiza completamente y entra en los microsomas. El péptido señal es removido una vez que la proteína ha ingresado a las vesículas.
Como resultado de estas y de otras experiencias, se ha podido diseñar la siguiente teoría: Cuando una proteína de secreción está siendo sintetizada por los ribosomas situados en el citosol, cuando ya van al menos 70 aminoácidos del extremo amino, se une la PRS. Por esta causa, la síntesis se detiene y sólo se reanuda cuando este complejo encuentra un receptor de la PRS, el cual está situado en las membranas del RER. La unión al receptor, libera la PRS y permite el anclaje del ribosoma a la membrana. Este paso es fundamental para la inserción cotransduccional de la proteína naciente en las cavidades del RER o en sus membranas. Algunos investigadores han acuñado el término TRANSLOCÓN para referirse al complejo de unión entre el ribosoma y las proteínas con las cuales interactúa en el RER.