Download

1 / 18
210 likes | 458 Views
RNA Binding Domains. James Polke, Institute of Neurology, London 10 th December 2009. RNA/Protein Interactions.

E N D
RNA Binding Domains James Polke, Institute of Neurology, London 10th December 2009
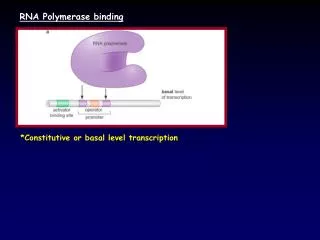
RNA/Protein Interactions • RNA is central to many cellular processes. It functions as a carrier of genetic information and as a catalytic and structural molecule. It is thought to have preceded DNA as the unit of inheritance and to have preceded proteins in catalysis. Most of these functions in the cell are now performed in combination with proteins. • Major RNA/Protein interactions • Transcription • RNA processing / transcriptional regulation • mRNA splicing/capping/polyadenylation • RNA transport • RNAi/miRNA • Translation • Ribosomal proteins (associating with rRNA and/or mRNA and/or tRNA) • RNAse enzymes • There are several classes of protein RNA binding domains that bind to RNA in many forms: • ssRNA / dsRNA / Other RNA structures (G-quartet etc.) • Sequence-specific binding or non sequence-specific • RNA binding, RNA/DNA binding etc..
Classes of Sequence-Specific ssRNA Binding Domains • RRM (RNA Recognition Motif) • Most abundant. Involved in most RNA post-transcriptional gene expression processes: mRNA and rRNA processing, RNA export and stability • KH Domain (hnRNP K Homology Domain) • 2nd most abundant. Involved in regulation of transcription, translation, splicing, and RNA transport and stability • Pumilio Homology Domain • Influence mRNA stability and translation by binding to 3'UTRs. • Zinc-Finger Domains • RNA stability
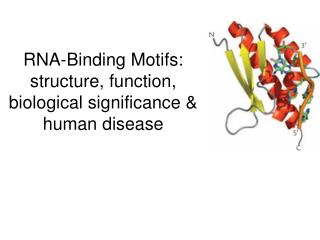
Clery, Blatter and Allain (2008) RNA Recognition Motif (RRM) • Also know as RNA Binding domain (RBD) is the most abundant RNA binding motif – present in 0.5-1.0% of ALL proteins. • Typically ~90aa. 4 x β-sheets, 2 x α-helices:
RNA Recognition Motif (RRM) • 2-8 bases of RNA typically binds to β-sheets (hydrogen bonding), • The RRM is composed of 2 core sequences: Ribonucleoprotein consensus sequences 1 and 2 (RNP-CS1 and 2), 8 and 5 amino acids, respectively. • Loops between sheets and helices can also bind RNA. Possibly a later evolutionary adaptation to expand the range of target RNAs. • It's recently been shown that some RRM domains can also bind proteins. • Some motifs are more flexible than others in their sequence specificity: Can bind to a range of less-conserved sequences. • Individual RRMs have a low binding affinity for RNA. Affinity and specificity are increased by multiple RRM motifs in many proteins • RRM domains can be predicted in proteins by homology with other motifs, but more research is required to accurately predict the conformation of domains and their sequence-binding specificity. Crystal structure provides the best information.
Messias and Sattler (2004) KH Domains • Highly abundant throughout archaea bacteria and eukaryotes • Binding site composed ~70 amino acids that form a fold between 3 x β-sheets, 3 x α-helices. KH1 and KH2 differ in the arrangement of these structures. • RNA binding affinity varies between individual proteins, reflecting the multiple roles of KH domains • Like RRM domains, proteins often have multiple KH domains
FMR-1: KH Domain mutation • Fragile X is usually caused by a lack of functional FMRP following CGG methylation and transcriptional silencing. However, the FMR-1 Ile304Asn point mutation causes extremely severe Fragile X. • Amino acid 304 is in the KH2 domain of FMRP. There was controversy over the effect of this mutation at the protein level, based on extrapolation of the known structure of other KH domains. But crystallisation of the FMRP KH1/2 complex revealed that it's in the hydrophobic core of the domain, destabilising the KH domain complex rather than affecting RNA binding itself.
Mutations in Other RNA Binding Genes • Mostly multi-system disorders • GARS (CMT2/dSMA-V): Glycyl-tRNA synthetase, involved in protein synthesis by catalyzing aminoacylation of glycyl-tRNA, which is required for binding tRNA with cognate amino acids. Pathology restricted to axons. • Diamond-blackfan anaemia: Anaemia, congenital malformations, growth retardation. AD. Caused by mutaitons one of 7 ribosomal proteins • Dyskeratosis congenita: A disease of Telomere shortening. Mutations in at least 3 ribonucleoproteins in the telomerase complex can cause this disease (can also be casued by mutationsin other telomerase complex proteins).
Mutations in Other RNA Binding Genes • Childhood ataxia with central nervous system hypomyelination: Symptoms include spasticity and ataxia, optic atrophy. Progressive. Caused by mutations in one of the 5 subunits that make up the translation initiation factor eIF2B. • Cartilage-hair hypoplasia: Short-limbed dwarfism. Skeletal abnormalities, immunological defects. Caused by mutions in RMRP, a nuclear-encoded mitochondrial ribonucleoprotein. • Wolcott-Rallison syndrome: infancy-onset diabetes mellitus and multiple epiphyseal dysplasia. Caused by mutations in EIF2AK3 (initiation factor 2-alpha kinase-3).
Pathology – Splicing • Mutations that affect mRNA splicing: • Disruption of constitutive splice sites • Activation of cryptic splice sites • Mutations in Intronic/Exonic splice Enhancers/Inhibitors • Mutations/variants can shift the balance of splice variants with potentially pathogenic effects: E.g. CTFR polyT in intron 8: 5T, 7T or 9T, the shorter the polyT the higher the likelihoood of aberrant splicing (skipping exon 9). - 5U/missense heterozygotes can have CBAVD. 5U/5U homozygotes can have CBAVD, though penetrance is modulated by a 2nd variable (polyUG) tract nearby. - Tissue-specific differences in splicing are thought to explain CBAVD – exon 9 exclusion is more likely in the vas deferens E.g. SMN2 exon 7 inclusion
Pathology – Disruption of Assembly of Functional Spliceosomes • At least 4 genes that cause AD Retinitis Pigmentosa encode proteins involved in spliceosome assembly: PRPF31, HPRP3, PRPC8, SNRNP200. • The most common cause of SMA is homozygous loss of the SMN1 gene, which is a regulator of spliceosome assembly • The above diseases are caused by cell-specific disruption of a ubiquitous process, how does it cause specific symptoms? • Splicing factors may be ubiquitous, but individual gene expression isn’t, particular introns/exons may have different sensitivities for splicing factors. • In SMA the defect may alter transport of specific RNAs along axons to the synapse of motor neurones
Pathology - Myotonic dystrophy • DM1: CTG expansion in 3'UTR of DMPK gene causing a multisystem disorder. Accumulation of (CUG)n RNA which adopt a hairpin 2° structure. • A majority of the pathology is due to trans effects of the mutant transcript: • Sequestration of TFs (e.g. SP1, STAT1/3, RARγ) with downstream effects on expression of multiple genes (e.g. CLCN1, Cl and K metabolism genes) • Sequestration of splicing factors: MBNL1-3 (Muscleblind like): Regulate splicing of Cardiac troponin and insulin receptor (IR) genes CUG-BP1: Activity of this alternate splicing factor is increased and it localises in the cytoplasm of myoblast, skeletal muscle and heart tissue, though it does not bind to CUG hairpins. Thought to antagonize the splicing activity of MBNL proteins hnRNP H: In normal cells hnRNP H complexes with CUG-BP1, inhibiting IR exon 11 inclusion – but this is countered by the activity of MBNL proteins. In the presence of the CUG repeat, sequestration of MBNL promotes IR exon 11 exclusion.
Pathology - Myotonic dystrophy • (CUG)n hairpins may be DICER targets, with two effects on expression: • Downregulating DMPK expression • Generating siRNAs that in turn act to downregulate the expression of other genes with complimentary trinucleotide repeats. • CUG-BP1 activates translation. In DM1 cells it is aberrantly phosphorylated at two residues with downstream effects on translation of several genes (p21, Mef 2A, C/EBPβ).
‘Toxic RNA’ effect in other disorders • SCA3: Li et al (2008) Nature453(19): 1107-1112 • FXTAS: Iwahashi et al (2006) Brain129: 256-271 • HDL-2 (Junctophilin 3): Rudnicki et al (2007) Annals of Neurology 61: 272-282
miRNAs • miRNAs downregulate gene expression by targeting homologous mRNAs for degradation, or by inhibiting translation • RNA binding proteins include DICER (dsRNA-binding RNAse) and several components of the RISC complex
miRNAs and Cancer • Alterations in the expression of miRNAs are now thought to be involved in a majority of human cancers. • There are several mechanisms: • Deletion/mutation of tumour suppressor miRNAs • Amplification/mutation of oncogenic miRNAs • miRNA dysregulation by transcription factors • miRNAs as targets/regulators of epigenetic changes Review: Croce (2009)
References Auweter SD, Oberstrass FC, Allain FH. Sequence-specific binding ofsingle-stranded RNA: is there a code for recognition? Nucleic Acids Res. 2006;34(17):4943-59. Epub 2006 Sep 18. Chen Y, Varani G. Protein families and RNA recognition. FEBS J. 2005May;272(9):2088-97. Cléry A, Blatter M, Allain FH. RNA recognition motifs: boring? Not quite. Curr Opin Struct Biol. 2008 Jun;18(3):290-8. Llamusí B, Artero R. Molecular Effects of the CTG Repeats in Mutant Dystrophia Myotonica Protein Kinase Gene. Curr Genomics. 2008 Dec;9(8):509-16. Faustino NA, Cooper TA. Pre-mRNA splicing and human disease. Genes Dev. 2003 Feb 15;17(4):419-37. Messias AC, Sattler M. Structural basis of single-stranded RNA recognition. Acc Chem Res. 2004 May;37(5):279-87. Valverde R, Edwards L, Regan L. Structure and function of KH domains. FEBS J. 2008 Jun;275(11):2712-26. Epub 2008 Apr 15. Croce CM. Causes and consequences of microRNA dysregulation in cancer. Nat Rev Genet. 2009 Oct;10(10):704-14.