Download

1 / 26
260 likes | 512 Views
Intracellular Cellular Communication by Second Messengers: How information is sent inside a cell. Chapter 14: 10/28 10/31. What are the types of hormone? How much hormone is produced? What is a second messenger and why do cells need them? What is signal amplification?

E N D
Intracellular Cellular Communication by Second Messengers: How information is sent inside a cell.Chapter 14: 10/28 10/31 • What are the types of hormone? • How much hormone is produced? • What is a second messenger and why do cells need them? • What is signal amplification? • How do steroid change cellular activity? • What are G-proteins? • How does epinephrine change cellular activity?
How do APs self propagate across a synapse? A Great Test Question!Show how an action potential reaches the synapse causing the release of an excitatory NT. Then show how the NT creates a local depolarization in the second cell by using ligand-gated channels. Then discuss why the voltage-gated channels in the second cell open and permit AP propagation.
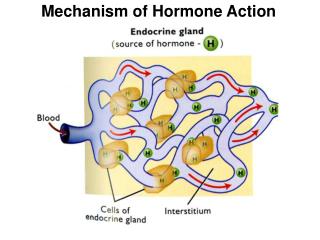
How are hormones (endocrine) classified with respect to site of production vs. site of cellular action? Distance to Target, Relative to Distance from Origin: 1) Endocrine: Carried from gland to body by blood! 2) Neuroendocrine: Carried from axon to body by blood! 3) Paracrine: local use by nearby cells only! 4) Autocrine: hormone effects cell that produced hormone! 5) Phermones: Carried outside body to other bodies H: often created in micro-nanogram/liter amounts! H: usually slower acting than neurotransmitters! H: often/secreted made when needed then destroyed! “Exocrine” secretions exit the body/do not enter blood!
Why are Second Messengers different from hormones? Why do hormones need Second Messengers? Second Messengers: deliver messages from outside the plasma membrane to intracellular targets that affect change in cell function. • 1) Can all hormones enter a cell? • Lipid soluble- • Water soluble- • Carriers required if not soluble! • 2) How much hormone is typically present in the fluid around the cell? • Micro to Nanogram/liter amounts • Hormones are rapidly degraded • 3) Why do we amplify the original message inside the cell? • 4) Ligand+Hormone changes an enzyme activity causing another molecule to be produced or its activity to me modified. • Second molecule directly changes activity of cell! • Hormones work by creating second messengers!
If a hormone (ligand) cannot enter the cytosol after bind a receptor, the ligand must create a second messenger within the cytosol to affect cellular change.
Second Messenger systems permit amplification of a weak hormonal signal in the cytosol creating large effects in the cell. They also permit excellent pathways for regulating (+ or -) intracellular effects.
No Second Messenger Required:Steroids and thyroxine are lipid soluble hormones requiring carriers for transport in the blood, but they can diffuse THROUGH the P.M. Once inside they find receptors (why) that carry then to specific “Hormone Response Elements” on the DNA modifying DNA transcription. This is how testosterone causes Actin/Myosin to be produced in muscle cell.
What are five classic second messenger systems that are used in cells to carry a message from the plasma membrane to its intracellular target? G-proteins to mediate effects of many but not all hormones 1) Phospholipase C create DAG and IP3 G-protein stimulation DAG Protein kinase C IP3 open calcium channels 2) Adenylyl Cyclases cyclic AMP G-protein stimulation Protein kinase A 3) Guanylyl Cyclase cyclic GMP Protein kinase G 4) Changes in intracellular calcium control kinases Secretion, contraction, enzyme regulation and more 5) Tyrosine Kinases phosphorylate proteins (No G-proteins)
G-Proteins: “trimers” (α+β+γ) that mediate many hormone signals in the cell.When the hormone is present, GTP is added to the alpha unit of the trimer. This allows the alpha-GTP to leave the trimer, diffuse to its target, and modify target activity. When GTP is hydrolyzed to GDP (takes a few seconds), the alpha subunit is inactivated, the system returns to the inactive state. Different types of Alpha subunit monomer can exist in a single cell or in different types of cell. GTPase activity of alpha subunitGDP + Pi converts the alpha subunit back to inactive state causing it to re-associate with the Beta and Gamma subunits
Phospholipase-C is activated by a G-alpha-q-GTP and mediates some effects of epinephrine among others. Ultimately IP3 and DAG activate other enzymes which in turn creates other down stream effects.
Adenylyl Cyclase produces the second messenger cAMP.AC is turned on by G-alpha-s-GTP stimulates ACAC is turned off by G-alpha-i-GTP inhibits ACcAMP removes the inhibitory units from a protein kinase which can phosphorylate serine/threonine residues on target proteins. PK-A
cAMP removes protein subunits that inhibit enzyme activity in the absence of the hormone in the target cell. Protein Kinase A PK-A Phosphorylates target serines How is cAMP degraded? Phosphdiesterase cuts cAMP open PDE inhibitors include caffeine Caffeine increase intracellular cAMP
How do these mechanisms work together? • A single cell can have hundreds of different receptors for different hormones. • A single cell can have many receptors activated at the same time. • The pathways of activation may amplify or cancel-out each others actions in the cell. • What a cell does is the sum of these pathways
Why does cytosolic calcium make a great signal for regulating the activity of cell enzymes? • Normal Intracellular and extracellular Ca++ levels: • 0.00001 mM 1.2 mM Ca++ Pumps are found on membranes at following spots: Plasma Membrane, Sarcoplasmic reticulum and E.R. • Three Regulated Ca++ Entry Methods: • 1) Ca++-VGC (PM): heart • 2) IP3 activated Ca++ channels (ER): fertilization • 3) Ca++ Activated (ryanodine) Ca-channels (SR): heart • This channel actually opens when Ca++ released from the PM binds to it! • This is a second source of Ca++ entry into the cytosol of cardiac myocyte • Ca++ Removal Methods: We have to get the signal out very fast! • Ca++-ATPase Ca++/Na+-Exchanger
Calcium is used in many ways, one of which is the creation of a message that fertilization has occurred. We a products of this!Ca++ influx is required for degranulation of cortisol granules (vesicles). Will fertilization occur in a calcium-free media?
Many of the intracellular effects of Calcium are due to a regulatory protein called calmodulin that is activated only by the PRESENCE OF calcium. • Ca++ needs to only reach 1 micromolar (10-6 M) for activation! • Ca++/Calmodulin Dependent protein kinases! • Ca++/Calmodulin activates nucleotide phosphodiesterases that break up cAMP, thereby limiting excess Ca++ entry into cell. • Ca++/Troponin dependent binding: required for removal of troponin and actin/myosin-contraction of skeletal/cardiac muscle!
Calmodulin finds and binds its intracellular target protein very efficiently when calcium is present! Ca++ binding causes a conformational change in its target (Calmodulin) that changes its activity in a cell.
The use of a calcium influx as a signal is a central dogma in cells.Cellular redundancy describes how several pathways can “converge” on the same target (influx of Ca++), this give a cell many ways to fine tune the physiological effect. Remember that Ca++ voltage gated channels can also rapidly increase intracellular calcium.
“GTP” can also be converted into cGMP, another important intracellular second messenger system! Viagra inhibits cGMP phosphodiestrase in the smooth muscle cells and prolongs the effect of cGMP resulting in extended vasodilation.
Hormones may bind a membrane receptor that changes internal protein activity by activating kinases/phosphatases On or Off Add or Remove Pi-groups to protein. • The insulin receptor is a tyrosine kinase that is turned ON by the presence of insulin (the ligand) at its binding site. • First: The receptor-ligand complex self-phosphorylates • Second: The Phosphorylated receptor phosphorylates OTHER proteins on the hydroxyle groups of their Tyrosine R-groups. • Third: Cascade leads to movement of glucse transporters from cytosol to plasma membrane so glucose enters sensitive cell from blood. • This causes Insulin-Sensitive cells (hepatocytes, adipocytes, and skeletal muscle cells to move glucose pores to the plasma membrane, this allows glucose to enter and leave the blood.
Tyrosine Kinase Family: There are many similar but slightly different types of tyrosine kinase that phosphorylate different yet “specific” tyrosines on specific intracellular proteins. Remember that the function of every kinase is balance by a phosphatase that removes Pi (reversal of effects).
How do growth factors work as tryosine kinases that mediate exocytosis, cell division, growth and sometimes cancer? Why do they often times serve as targets for chemotherapy? Exocytosis?? CANCER???
For Test: review these second messenger systemsBe able to describe each and the things that make it unique! • What are the hormone classes? • Structural vs. Site of Effect/origin • Phospholipase-C System: DAG/IP3 • G-Proteins: alpha, beta and gamma/inhibitor and stimulatory • Presence of intracellular calcium: how is it pumped in/out/stored? • Adenylyl cyclase: products and regulation? • How does cAMP affect cellular activity? • What does phosphodiesterase do to cAMP or cGMP? • Guanylyl cyclase: products and regulation • Calmodulin/troponin: what does it do when Ca++ is present? • How do tyrosine kinases work? • What are the amino acids targeted by kinases?
Indicate the function of each second messenger system: Be specific! What does each one do! 1 Phosphodiesterase 2 Calmodulin 3 Adenylyl cyclase 4 G-protein Trimer 5 Endoplasmic reticulum 6 Stimulatory α-GTP 7 Phospholipse-C 8 Phosphokinase-C (Protein Kinase C) 9 Calcium activated-Ca++ Channel 10 IP3-Activated Ca++ Channel 11) Tryosine kinase family