Download
1 / 1
10 likes | 75 Views
Explore the changes in glycinergic inhibition in the spinal cord during postnatal development in rats, impacting nociceptive circuits and sensory input transmission.

E N D
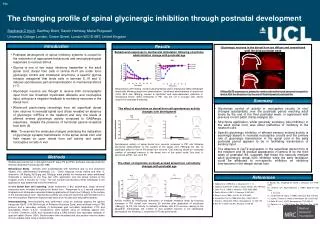
514 The changing profile of spinal glycinergic inhibition through postnatal development Stephanie C Koch, Geoffrey Brent, Gareth Hathway, Maria Fitzgerald University College London, Gower Street, London WC1E 6BT, United Kingdom Introduction Results Glycinergic neurons in the dorsal horn are diffuse and unrestricted until the third postnatal week Behavioural responses to mechanical stimulation following strychnine administration change with postnatal age • Postnatal development of spinal inhibitory systems is crucial for the maturation of appropriate behavioural and neurophysiological responses to noxious stimuli • Glycine is one of two major inhibitory transmitter in the adult spinal cord: dorsal horn cells in lamina III-VI are under tonic glycinergic control and intrathecal strychnine, a specific glycine receptor antagonist that binds cells in laminae II, III and V, induces spontaneous pain and sensitization to mechanical stimuli (1-7). • Glycinergic neurons are thought to receive both monosynaptic input from low threshold myelinated afferents and nociceptive input, acting as a negative feedback to excitatory neurones in the dorsal horn. • Whole-cell patch-clamp recordings from rat superficial dorsal horn neurons in neonatal spinal cord slices revealed an absence of glycinergic mIPSCs in the newborn and very low levels of afferent evoked glycinergic activity compared to GABAergic responses, despite the presence of functional glycine receptors from birth (9) • Aim: To examine the molecular changes underlying the maturation of glycinergic synaptic transmission in the spinal dorsal horn and their impact on upon dorsal horn cell activity and spinal nociceptive circuits in vivo P3 P10 I II III IV V VI I II III IV V VI P21 P40 I II III IV V VI Responses to vFh testing reveal a developmental shift in mechanical reflex withdrawal thresholds following strychnine administration. Intrathecal administration of strychnine (16.5, 82.5 and 165ng/g) causes a significant and dose-dependent reduction in mechanical flexion reflex thresholds in P21 rats but not in younger rats, where strychnine increases thresholds. • Diffuse GlyT2 expression is present in newborn dorsal horn and becomes focussed to lamina III of the dorsal horn by the end of the third week of postnatal life. Summary The effect of strychnine on dorsal horn cell spontaneous activity changes over development • Glycinergic control of activity in nociceptive circuits ‘in vivo’ changes substantially over the postnatal period reaching adult levels by the end of the third postnatal week, in agreement with previous in vitro patch clamp analysis (9). • Strychnine application, whilst generally excitatory (dis-inhibitory) in the adult spinal cord, was either ineffective or inhibitory in the newborn cord. • Specific glycinergic inhibition of afferent sensory evoked activity is seemingly absent in neonatal nociceptive circuits and the primary role of glycinergic transmission in the spinal cord in the early postnatal period appears to be in facilitating transmission of sensory input. • The absence of GlyT2 expression in the superficial dorsal horn of the newborn and its gradual appearance in lamina III by the third week of postnatal life, suggests that these the neurons mediate adult glycinergic dorsal horn inhibition while the early facilitation could be attributed to non-specific inhibition on inhibitory interneurons in the deeper dorsal horn. P21 P3 saline saline strychnine strychnine Spontaneous activity of spinal dorsal horn neurons increases in P21 rats following strychnine administration to the surface of the spinal cord (165ng/g) but has no significant effect on that of P3 dorsal horn neurons. Cfos staining shows an increased number of activated neurons in the dorsal horn of the spinal cords 2hrs post i.t. strychnine at P21, but not at P3. Methods Studies were carried out in rats aged 3 and 21 days (P3 and P21) and were licensed under the Animals (Scientific Procedures) Act 1986. Behavioural Study: Animals were anaesthetised with halothane and O2 and strychnine (Sigma U.K.) administered intrathecally (i.t.). Dose response curves before and after i.t. strychnine (16.5ng/kg, 82.5ng/g and 165ng/g) were plotted for mechanical reflex withdrawal thresholds in response to von Frey hair (vFh) application onto the dorsal surface of the hindpaw, every 5 minutes for 1 hour. The hair number that elicited a reflex withdrawal in 2/5 applications was determined to be the threshold. In vivo dorsal horn cell recording: Under isofluorane (1.8%) anaesthesia, singleneuronal responses were recorded throughout the dorsal horn. Responses to a 2 second cutaneous hindpaw brush stroke were recorded following application of strychnine (165ng/g) to the surface of the exposed spinal cord. Spontaneous activity pre and post strychnine was recorded over a one minute period. Maximum response was seen at 20 minutes post-drug administration. Immunostaining: Immunostaining was performed using an antibody against the glycine transporter GlyT2 (1:40,000 kind gift of Professor Francisco Zafra) and visualized using a TSA protocol and a secondary antibody of biotinylated goat anti-rabbit (Vectastain, FITC and secondaries Vector, U.S.A.). Cryosections were stained using an antibody against cfos (1:10,000; Chemicon, USA) and visualised using a DAB protocol and secondary antibody of goat anti-rabbit (Vector, USA). Sections were then visualised and cfos positive neurons drawn onto an acetate sheet using a camera lucida. The effect of strychnine on brush-evoked dorsal horn cell activity changes with postnatal age ipsi contra ipsi contra P3 P21 References 9. Baccei ML, Fitzgerald M (2004) J Neurosci. 24: 4749-4757. 10. Ottersen OP, Storm-Mathisen J (1987) Biochem Soc Trans. 2: 210-213. 11. van der Pol AN, Gorcs T (1988) J Neurosci 8(2) 472-492. 12. Jursky F, Nelson N (1995) J Neurochem. 64: 1026-1033. 13. Spike RC et al. (1997) Neuroscience 77(2):543-51. 14. Poyatos I et al. (1997) Brain Res Mol Brain Res. 49:63-70. 1. Todd AJ et al (1995) Eur. J. Neurosci. (7): 1-11. 2. Todd AJ, Sullivan AC (1990) J. Comp. Neurol. 296: 496-505. 3. Zafra F et al. (1995) J. Neurosci. 15(5): 3952-3969. 4. Zarbin MA et al (198 1) J. Neurosci. 5: 532-547. 5. Beyer C et al (1985). Life Sci. 37: 875–882. 6. Campistron G et al (1986) Brain Res. 376(2):400-5. 7. Sivilotti L, Woolf CW (1994) J Neurophysiol. 72:169-179. 8.Ben-Ari Y (2002) Nat Rev Neurosci. 9:728-39. saline saline strychnine strychnine Activity evoked by mechanical stimulation of hindpaw receptive fields by brushing increases in P21 dorsal horn neurons 20 minutes after application of strychnine (165ng/g). At P3, this activity is markedly inhibited, with 3/14 neurons ceasing to fire altogether. Interestingly, the number of cfos postitive neurons is not significantly decreased 2 hrs following i.t. strychnine in P3 rat spinal cords.