Download

1 / 25
250 likes | 454 Views
Monoklony in planta. PLANTIBODIES. Cechy produkcji monoklonów w roślinach. Przewiduje się wprowadzenie 25-50 nowych terapeutycznych mAb w ciągu 5 lat. Wymaga to potrojenia światowych zasobów bioreaktorów. Nie ma na to pieniędzy ani czasu.

E N D
Monoklony in planta PLANTIBODIES
Cechy produkcji monoklonów w roślinach • Przewiduje się wprowadzenie 25-50 nowych terapeutycznych mAb w ciągu 5 lat. Wymaga to potrojenia światowych zasobów bioreaktorów. Nie ma na to pieniędzy ani czasu. • Do produkcji masowej w roślinach nie trzeba inwestować w bioreaktory (redukcja wkładu pieniędzy o 90%). • Rośliny posiadają zadziwiająco podobny do zwierząt aparat biosyntezy białka • Pozwalają uniknąć zakażeń odzwierzęcych (wirusy, priony, toksyny). Dodatkowo znikają koszty kontroli zakażeń i ich zwalczania
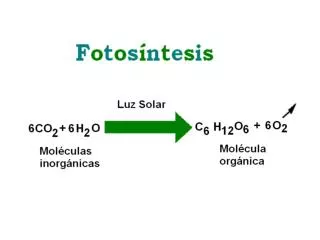
Najefektywniejsze bioreaktory-molecular farming • Rośliny to najefektywniejsi na świecie producenci białka. Najniższe koszty utrzymania hodowli dostępna energia
Uzyskiwanie cDNA dla przeciwciał • Z biblioteki fagowej (phage display) poprzez izolację DNA z fagów prezentujących na swojej powierzchni pożądane przeciwciało • Bezpośrednio z kolonii hybrydom poprzez izolację ich RNA i odwrotną transkrypcję na cDNA. • Następnie otrzymanym cDNA transformuje się roślinę wybraną metodą.
Porównanie • 1-akrowe (63m * 63m) poletko kukurydzy daje tyle samo przeciwciał co 100,000 litrów komórek (ponad 6 ogromnych bioreaktorów)czyliok. 18 kg przeciwciał na rok. • Plantibodies mogą kosztować na rynku $0,1 - 1$ za gram, te z kultur komórkowych kosztują teraz $200 - $650 za gram
Koszty produkcji białek w różnych systemach Stosunek cenowy 1:3000
Porównanie czasowe Ale nie można w nich produkować mAb lata
Promotory dla genów przeciwciał • Promotory tkankowo-specyficzne pozwalają wybrać pożądane miejsce ekspresji (liście, korzenie, nasiona...). Ułatwia to oczyszczanie przeciwciał z rośliny i uzyskanie odpowiedniej ich akumulacji • Zwraca się także uwagę na bezpieczeństwo ekologiczne. Np. pomidorowy promotor (HMGR2) z genu aktywowanego szokiem mechanicznym (MeGA). Ekspresja transgenu jest aktywowana dopiero w momencie zbierania i przetwarzania rośliny co prowadzi do błyskawicznej translacji białka (24h)
Oczyszczanie mAb z roślin • Akumulacja w ciałkach tłuszczowych nasion Transgeniczna oleozyna wchodząca w skład ich powierzchi zawiera w sobie białko A wiążące Ab Faza tłuszczowa Faza wodna
Oczyszczanie – c.d. Nie trzeba niszczyć roślin w celu zebrania przeciwciał • Skierowanie białka do ryzosekrecji umożliwia jego odzysk wprost z podłoża hydroponicznego. • Skierowanie do wydzieliny gutacyjnej- odzysk wprost z powierzchni liścia
Ekspresja w nasionach kukurydzy • Białko w nasionach można długo składować (3 lata w lodówce) i jest zawarte w małej objętości co ułatwia oczyszczanie
Otrzymano już wszystko wszystko fragment SC ramię J
secretory-IgA (wydzielnicze) • Wykorzystuje się je (w dużej ilości – kilka gramów na dzień) gdy przeciwciała muszą być stosowane miejscowo lub wziewnie na błony śluzowe. (Zapobieganie infekcjom patogenów wenerycznych, astma) • Są odporne na proteolityczne środowisko nabłonków • Są bardziej zachłanne (dimery) koagulacja bakterii (zatem dodatkowa ochrona niespecyficzna) • U ssaków SC jest „dokładany” do IgA w komórkach nabłonka podczas sekrecji do światła narządu (jelito, przewody oddechowe). Rośliny transfekowane genami dla IgA i S.C. (=komponentu wydzielniczego) potrafią złożyć sIgA od razu, w jednej komórce.
Powstają różne konstrukty białkowejednak ilość sIgA jest zadowalająca
Próchnica zębów, o której było głośnoPrzykład produkcji sIgA w roślinach • Immunizując myszy otrzymano przeciwciała IgG przeciwko białkom powierzchniowym ludzkiego Streptococcus mutans (powodują próchnicę) • Konstrukty hybrydowych przeciwciał sIgA/G (z sIgA - wiązanie przez SC, zdolność dimeryzacji, z IgG – część zmienna) wklonowano do tytoniu. Ekspresja białka w liściach. Aplikowano ludziom doustnie wyizolowane przeciwciała. • Są tak samo powinowate jak oryginały IgG z myszy, lecz bardziej zachłanne! (tworzą dimery) • Bardziej odporne na proteolizę przez bakterie • Brak rekolonizacji przez bakterie w przeciągu 3 miesięcy (tyle trwał eksperyment)
Weterynaria - Drób • Jako pasywna i aktywna szczepionka przeciwbakteryjna w paszy-duża ilość Ab, nie trzeba oczyszczać • Pozwalają uniknąć stosowania antybiotyków które potem zjadamy • Ziarna kukurydzy zawierają kurze sIgA, które w przewodzie pokarmowym zwiększają wyłapywanie patogenu i jego prezentację komórkom ukł. immunologicznego • Wytwarza się systemowa odporność swoista
Widmoglikozylacji • Przeciwciała to glikoproteiny. IgG mają ok. 30 N-glikozylacji. Np. na każdej z 2 domen CH2 regionu stałego (Fc) niezbędna jest ona do wywołania toksyczności przeciwciało-zależnej ADCC (terapia raka)
Glikoinżynieria • Oczywiście glikozylacja jest inna niż u zwierząt. Rośliny np. wykorzystują wiązania (1–3)-fukoza oraz(1–2)- ksyloza, niespotykane u ssaków. Brak natomiast kwasu sjalowego (całego szlaku syntezy) • U hybrydom i myszy jest też inna niż u ludzi. Ludzkie Ab mają tylko N-acetyloneuraminowy (NANA), gryzonie mieszaninę NANA + N-glikozylo-neuraminowego. • Nie zaobserwowano wpływu toksycznego czy immunogennego ani zmniejszonego powinowactwa z tego powodu. • Jednak jest to potencjalny problem, zwłaszcza gdy dla swoistości Ig wymagana jest glikozylacja lub ważny jest fragment Fc aktywujący makrofagi i układ dopełniacza.
Glikoinżynieria DODAĆ USUNĄĆ
Glikoinżynieria • Każda reakcja przyłączenia cząsteczki cukru jest prowadzona przez specyficzną transferazę( N-oligo-saccharyltransferase, galactosyltransferase, sialyltransferase), rozpoznającą (w ER lub GA) strukturę przestrzenną fragmentu białka które modyfikują.
Glikoinżynieria-rozwiązania • Poznano zestaw genów jakie należy dodać, wyciszyć lub wzmocnić aby uzyskać glikozylację całkowicie ludzką. • Można uzyskać ich ekspresję w roślinie lub transgenicznych gryzoniach • Możliwa także modyfikacja enzymami in vitro pooczyszczeniu białka, ale jest to kosztowne
Glikoinżynieria Wprowadzenie transgenu ssaczej transferazy
Glikozylacja DODAĆ cukry USUNĄĆ cukry In vitro Transgeny dla glikotransferaz Retencja w ER (KDEL), nie ma dalszej glikozylacji w GA Knock-out genu transferazy
Zastosowania plantibodies • Cele medyczne i weterynaryjne (rak, HIV, HCV, próchnica, leczenie przedawkowań...) • Pasywna immunizacja roślin przeciwko grzybom, wirusom, insektom (produkcja np. na liściach) • Wydzielanie przez rośliny do ziemi abzymów rozkładających np. mocznik hamujący fotosyntezę • Immunomodulacja roślin i zwierząt-poznawanie funkcji blokowanych przeciwciałem białek • Antitransformation-kierowanie genów np. śmierci do konkretnej komórki
1. Anonymous (2002) Wkly. Epidemiol. Rec. 77, 109–120. • 2. Wilde, H., Tipkong, P. & Khawplod, P. (1999) J. Travel Med. 6, 238–242. • 3. Daniell, H., Streatfield, S. J. & Wycoff, K. (2001) Trends Plant Sci. 6, 219–226. • 4. Hiatt, A., Caffertey, R. & Bowdish, K. (1989) Nature 342, 76–78. • 5. Ma, J. K., Hikmat, B. Y., Wycoff, K., Vine, N. D., Chargelegue, D., Yu, L., • Hein, M. B. & Lehner, T. (1998) Nat. Med. 4, 601–606. • 6. Zeitlin, L., Olmsted, S. S., Moench, T. R., Co, M. S., Martinell, B. J., Paradkar, • V. M., Russell, D. R., Queen, C., Cone, R. A. & Whaley, K. J. (1998) Nat. • Biotechnol. 16, 1361–1364. • 7. Khoudi, H., Laberge, S., Ferullo, J. M., Bazin, R., Darveau, A., Castonguay, • Y., Allard, G., Lemieux, R. & Vezina, L. P. (1999) Biotechnol. Bioeng. 64, • 135–143. • 8. Bouquin, T., Thomsen, M., Nielsen, L. K., Green, T. H., Mundy, J. & Dziegie, • M. H. (2002) Transgenic Res. 11, 115–122. • 9. Conrad, U. & Fiedler, U. (1998) Plant Mol. Biol. 38, 101–109. • 10. Mann, M. & Jensen, O. N. (2003) Nat. Biotechnol. 21, 255–261. • 11. Faye, L., Gomord, V., Fitchette-Laine, A. C. & Chrispeels, M. J. (1993) Anal. • Biochem. 209, 104–108. • 12. van Ree, R., Cabanes-Macheteau, M., Akkerdaas, J., Milazzo, J. P., Loutelier- • Bourhis, C., Rayon, C., Villalba, M., Koppelman, S., Aalberse, R., Rodriguez, • R., et al. (2000) J. Biol. Chem. 275, 11451–11458. • 13. Helenius, A. & Aebi, M. (2001) Science 291, 2364–2369. • 14. Navazio, L., Miuzzo, M., Royle, L., Baldan, B., Varotto, S., Merry, A. H., • Harvey, D. J., Dwek, R. A., Rudd, P. M. & Mariani, P. (2002) Biochemistry 41, • 14141–14149. • 15. Henderson, J., Bauly, J. M., Ashford, D. A., Oliver, S. C., Hawes, C. R., • Lazarus, C. M., Venis, M. A. & Napier, R. M. (1997) Planta 202, 313–323. • 16. Bauly, J. M., Sealy, I. M., Macdonald, H., Brearley, J., Droge, S., Hillmer, S., • Robinson, D. G., Venis, M. A., Blatt, M. R., Lazarus, C. M. & Napier, R. M. • (2000) Plant Physiol. 124, 1229–1238. • 17. Sharp, J. M. & Doran, P. M. (2001) Biotechnol. Bioeng. 73, 338–346. • 18. Rudd, P. M., Elliott, T., Cresswell, P., Wilson, I. A. & Dwek, R. A. (2001) • Science 291, 2370–2376. • 19. Dietzschold, B., Gore, M., Casali, P., Ueki, Y., Rupprecht, C. E., Notkins, A. L. • & Koprowski, H. (1990) J. Virol. 64, 3087–3090. • 20. Prosniak, M., Faber, M., Hanlon, C. A., Rupprecht, C. E., Hooper, D. C. & • Dietzschold, B. (2003) J. Infect. Dis. 187, 30386–30389. • 21. Datla, R. S. S., Bekkaoui, F., Hammerlindl, J. K., Pilate, G., Dunstan, D. I. & • Crosby, W. L. (1993) Plant Sci. 94, 139–149. • 22. Kay, R., Chan, A., Daly, M. & MacPherson, J. (1987) Science 236, 1299–1302. • 23. Ko, K., Norelli, J. L., Reynoird, J.-P., Boresjza-Wysocka, E., Brown, S. K. & • Aldwinckle, H. S. (2000) Biotechnol. Lett. 22, 373–381. koniec