Download
1 / 62
630 likes | 1.12k Views
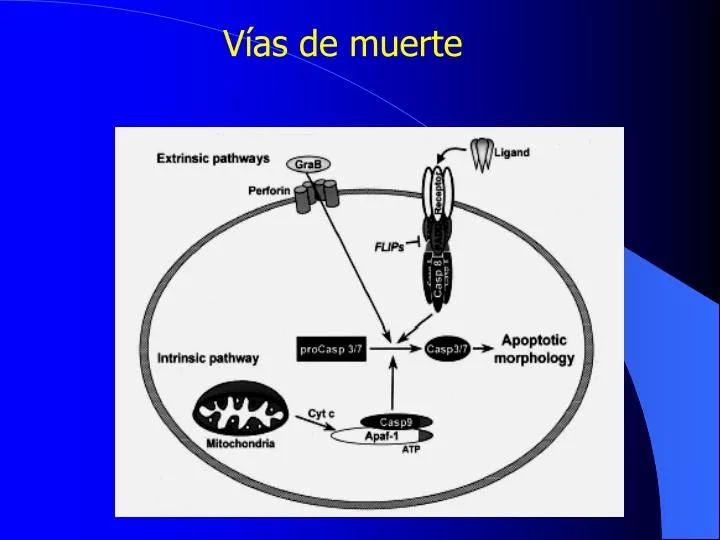
Vías de muerte. caspase c ysteine-dependent asp artate specific prote ase Especificidad dominante para sustratos conteniendo Asp (específico de caspasas) Excepción : Granzima B: activador de caspasas Contienen un pentapéptido conservado: QACXG

E N D
caspase • cysteine-dependentaspartate specific protease • Especificidad dominante para sustratos conteniendo Asp (específico de caspasas) Excepción: Granzima B: activador de caspasas • Contienen un pentapéptido conservado: QACXG conteniendo la cisteína del sitio activo(común en cisteína proteasas) • Horvitz y cols.: Caenorhabditis elegans • CED 3 CED4 CED9
CED 3: proteína pro-apoptótica con homología de secuencia con: interleukin-1-converting enzyme (ICE)-like proteasas pro-IL-1 IL-1 por clivaje entre Asp 116-Ala 117. Para unificar la nomenclatura: CASPASAS Caspasa 1: primera caspasa identificada (la numeración de los aácidos de todas las caspasas se refiere a ella)
Procesamiento de caspasas • * Residuos esenciales para la catálisis • Residuos para reconocimiento de sustrato
Interrogantes acerca de la función de algunas caspasas * Caspasa 2: iniciadora vía TNFR * Caspasas 4,5 y 13: rol desconocido Se las agrupa como activadoras de Citoquinas por su similitud con la secuencia con caspasa 1 Pero: caspasa 13 es activada por caspasa 8?? Significado fisiológico desconocido
Sustratos de caspasas • Caspasas activ. de citoquinas (1, 4 y 5):(W/Y)EHD • Caspasas iniciadoras (8, 9 y 10): (I/L/V)EHD • Caspasas ejecutoras (3 y 7) y caspasa 2: DEXD • Uso de sustratos selectivos en mezclas de caspasas • Es complicado • La actividad y abundancia de cada caspasa, determina el resultado. La estructura ternaria es relevante
Sustratos de Caspasas Elementos del citoesqueleto: actina, fodrina, proteína tau y catenina Enzimas encargadas de reparar (PARP) o degradar (DNAasa) el DNA Factores de transcripción: Rb, MDM2 Proteínas quinasa y fosfatasas: PKC, fosfatasas 2-alfa, Akt Productos de la familia Bcl-2 CAD-ICAD
Estructura de las caspasas Heterodímeros: cada uno contiene un sitio catalítico compuesto por residuos de ambas subunidades Cada sitio activo contiene un S1 (+)(altamente conservado) para unir el Aspartato P1 (-) del sustrato P2 y P3 tienen un efecto limitado sobre el clivaje del sustrato P4 y S4 son relevantes en la especificidad por sustrato
Sustratos sintéticos Secuencia tetrapeptídica de preferencia de caspasas Sustratos sintéticos: tetrapéptidos conjugados a: 1) un cromóforo pNA (espectrofotometría) 2) un fluorocromo: AMC (7-amino-4-metilcumarina) AFC (7-amino-4- trifluorometil- cumarina) (espectrofluorometría)
La secuencia tetrapeptídica es una secuencia de “preferencia” y no absoluta Muchas caspasas pueden clivar el mismo sustrato “in vitro” aunque con distinta eficiencia. La elección depende de la concentración, tiempo de reacción y accesibilidad
Inhibidores Sintéticos Se unen al sitio activo de las caspasas en forma reversible o irreversible Algunos forman uniones covalentes con la cisteína del sitio activo Bloquean o retrasan la muerte Pueden ser tóxicos para la célula (incub. t largos) Como ocurre con los sustratos de caspasas, los inhibidores no son específicos para una caspasa
Otros roles de las caspasas • Regulación de la diferenciación celular: FADD es un regulador clave del desarrollo de células T • Proliferación celular ( de células T) y progresión del ciclo celular • Motilidad celular • Internalización de receptores (FAS R)
Mecanismos de activación de caspasas Homoactivación: requiere del reclutamiento de zimógenos a proteínas adaptadoras: caspasas iniciadoras 8 y 10: DISC, mediado por R de muerte caspasa iniciadora 9: Apoptosoma, compuesto por Apaf1, citocromo c, dATP/ATP y caspasa 9 Heteroactivación: ocurre por acción de otras proteínas sobre los zimógenos caspasas
Dominios de Apaf-1 Card: se une al Card de caspasa 9 NBD: dominio de unión a nucleótidos, involucrado en la oligomerización WD40: involucrados en interacciones proteína- proteína
* El dominio CARD no está normalmente expuesto y no se une a procaspasa 9 a menos que Apaf1 sea activado por dATP y citocromo c * El acceso al dominio CARD está normalmente bloqueado por las WD40 * ATP y cit.c hacen que Apaf-1 sufra un cambio con- formacional que expone el dominio CARD uniendo luego la pro-caspasa 9
Apoptosoma: la rueda de la muerte El sitio de unión para Cit. c con Apaf-1 no ha sido caracterizado WD-40 regulan el sitio de unión La unión de Cit. C expone los sitios de unión para dATP
La homodimerización parece ser la clave para la activación de la procaspasa 9 Un evento único es que sólo uno de los dos polipép- tidos se vuelve activo Probablemente los monómeros de procaspasa 9 unidos al apoptosoma recluten nuevos monómeros de procaspasa 9 para formar la holoenzima El apoptosoma permitiría la interacción antiparalela de dos monómeros inactivos para formar el dímero asimétrico activo
Interacciones caspasa 9-apoptosoma Caspasa 9 libre: actividad mínima el apoptosoma actúa como holoenzima La actividad del zimógeno y la forma procesada aumentan 2000 veces por unión al apoptosoma Estequiometría Apaf-1/caspasa 9: 1:1 El procesamiento proteolítico en el linker no es necesario para la activación de caspasa 9 El dominio N-terminal no se corta, la caspasa puede permanecer asociada al apoptosoma y activar casp. ejecutoras
Niveles de regulación de las caspasas • Hay precursores de caspasas expresados en forma • constitutiva deben estar estrechamente • reguladas • Los precursores de caspasas están presentes en • una conformación que previene la autocatálisis • 2) Necesitan de proteínas adaptadoras y cofactores • que cambien su conformación y las vuelvan activas • 3) Compartimentalización de caspasas y cofactores
Niveles de regulación de caspasas • Unión específica de inhibidores naturales: • FLIP • IAPs • A diferencia de la familia Bcl-2, IAPs bloquean • tanto la vía mitocondrial como la del R por unión a • caspasas iniciadoras y efectoras
Localización de las caspasas * Normalmente citosólica * También se han encontrado en: núcleo RE mitocondria Algunas caspasas pueden traslocarse de un compar- timiento celular a otro en presencia de un estímulo apoptótico Caspasas 2 y 9: de la mitocondria al citosol Caspasa 7: del citosol a la mitocondria
Miembros de la familia IAP • Para oponerse a la destrucción celular mediada por • caspasas se cuenta con dos clases de inhibidores • * Miembros de la familia Bcl-2: bloquean la vía mitocon- • drial • * Miembros de la familia IAP: bloquean la vía mitocon- • drial y la vía del receptor • Se unen a caspasas iniciadoras y efectoras e inhiben • su actividad
Miembros de la familia IAP NIAP:atrofia muscular espinal BIR: Cys/His RING, UBC: ubiquitin ligasa Degradación de caspasas unidas vía proteasoma
XIAP Es el más potente inhibidor de caspasas 3, 7 y 9 Aún no se conoce la función del dominio BIR 1 La región que circunda BIR 2 une específicamente caspasas 3 y 7 BIR 3 une caspasa 9 BIR 2 no tiene rol directo en la inhibición de caspa- sas
BIR3 de XIAP se une a la caspasa 9 clivada BIR 3 reconoce un tetra- péptido Ala-Thr-Pro-Phe Este tetrapéptido también se encuentra en SMAC/DIABLO
Posibles funciones de BIR 1 * Puede actuar estabilizando XIAP * Puede interactuar e inhibir caspasas aún no identificadas * Puede proveer una interfase para la unión de proteínas que interactúan con XIAP, modificando su actividad
¿Porqué tener una proteína con diferentes dominios que inhiban distintas caspasas? Permite la regulación simultánea de varias caspasas XIAP se une al apoptosoma donde interactúa con caspasas 9 y 3 sugiriendo que previene: * la activación de caspasa 3 por caspasa 9 * la liberación de la caspasa 3 del apoptosoma
XIAP se asocia con el apoptosoma La proximidad de XIAP a caspasas 3 y 9 y el hecho de que distintos dominios inhiban caspasas 3 y 9 justificarían la alta eficiencia y potente inhibición de estas caspasas por XIAP
Inhibiendo a los inhibidores SMAC/DIABLO Second Mitochondria derived Activator of Caspase Conecta a los IAPs con la MC controlada por la M Se sintetiza como un precursor que se trasloca a la mitocondria Allí se remueve un N-term de 55 aácidos (sec. de targeting) y se expone un tetrapéptido Ala-Val-Pro-Ile esencial para su unión a IAPs Interactúa con: XIAP, c-IAP ½ y survivina
Activación de caspasas por SMAC/DIABLO El linker NH2-t de BIR2 ocupa el sitio de unión del sustrato de caspasas
XAF1: una proteína de interacción con XIAP XIAP- Asociator Factor I Antagoniza la habilidad de XIAP para inhibir la actividad de caspasas “in vitro” A diferencia de SMAC/DIABLO no necesita ser procesado Está localizado en el núcleo y no se conoce el mecanismo de inhibición. Se propone que ocurre en el núcleo
Los dos mecanismos posibles de regulación de XIAP Smac/Diablo: No tendría efecto Sobre la vía del Receptor XAF-1: su locali- zación nuclear sugiere que puede interceptar ambas vías apoptóticas
Proteínas anti-apoptóticas • BH1-BH4: son indispensables para su actividad de • supervivencia • BH1-BH4: no tienen actividad catalítica pero intervie- • nen en la interacción con otras proteínas • BH1-BH3: forman un surco hidrofóbico en la parte • funcional; BH4 interviene en la estabilización del bol- • sillo hidrofóbico • Si se remueve BH4 la proteína se transforma en pro- • apoptótica
Miembros pro-apoptóticos * Ante un estímulo apoptótico exponen su dominio BH3 mediante un cambio conformacional (Bcl-2 no puede hacerlo) * Este dominio BH3 es el que va a interaccionar con el bolsillo hidrofóbico de las proteínas anti-apoptóticas * El cambio conformacional puede ocurrir por: defosforilación: Bad clivaje: Bid * La consecuencia del cambio conformacional es la traslocación a la mitocondria
Bax: targeting, inserción, oligomerización y formación del canal No hay canal canal
Modo de acción de Bcl-2-like y Bax-like factors
El casposoma de C.Elegans Modo de acción de una proteína BH3-only BH3-only Sensores y Mediadores de las res- puestas apoptóticas
Targeting de Bid y Bad a la mitocondria (A): t-Bid se puede unir a Bax o Bcl-Xl (B): independiente de caspasas (C): un aumento en la conc. de Ca++ induce la calcineurina fosfatasa que defosforila Bad






















