Download

1 / 18
180 likes | 333 Views
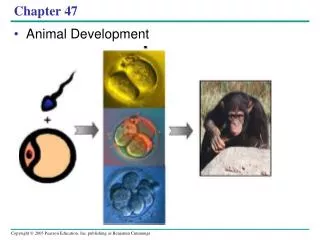
Ch. 47. THE STAGES OF EARLY EMBRYONIC DEVELOPMENT. From egg to organism, an animal’s form develops gradually: the concept of epigenesis. An embryo is not preformed in an egg; it develops by epigenesis, the gradual, gene-directed acquisition of form.

E N D
From egg to organism, an animal’s form develops gradually: the concept of epigenesis • An embryo is not preformed in an egg; it develops by epigenesis, the gradual, gene-directed acquisition of form.
Fertilization activates the egg and brings together the nuclei of sperm and egg • Fertilization both reinstates diploidy and activates the egg to begin a chain of metabolic reactions that triggers the onset of embryonic development. • The acrosomal reaction, which occurs when the sperm meets the egg, releases hydrolytic enzymes that digest through material surrounding the egg. • Gamete fusion depolarizes the egg cell membrane and sets up a fast block to polyspermy.
Sperm-egg fusion also initiates the cortical reaction, involving a signal-transduction pathway in which calcium ions stimulate cortical granules to erect a fertilization envelope that functions as a slow block to polyspermy. • In mammalian fertilization, the cortical reaction hardens the zona pellucida as a slow block to polyspermy. • SEE Figure 47.5 on page 1002
Cleavage partitions the zygote into many smaller cells • Fertilization is followed by cleavage, a period of rapid cell division without growth, which results in the production of a large number of cells called blastomeres. • Holoblastic cleavage, or division of the entire egg, occurs in species whose eggs have little or moderate amounts of yolk. • Meroblastic cleavage, incomplete division of the egg, occurs in species with yolk-rich eggs. • Cleavage planes usually follow a specific pattern relative to the animal and vegetal poles of the zygote. • In many species, cleavage creates a multicellular ball called the blastula, which contains a fluid-filled cavity, the blastocoel. • SEE FIGURE 47.8
Gastrulation rearranges the blastula to form a three-layered embryo with a primitive gut • Gastrulation transforms the blastula into a gastrula, which has a rudimentary digestive cavity (the archenteron) and three embryonic germ layers: the ectoderm, endoderm, and mesoderm. • SEE FIGURE47.10 p.1006
In organogenesis, the organs of the animal body form from the three embryonic germ • Early events in organogenesis in vertebrates include formation of the notochord by condensation of dorsal mesoderm, development of the neural tube from folding of the ectodermal neural plate, and formation of the coelom from splitting of lateral mesoderm. • SEE FIGURE 47.11, p. 1008
Amniote embryos develop in a fluid-filled sac within a shell or uterus • Meroblastic cleavage in the yolk-rich, shelled eggs of birds and reptiles is restricted to a small disc of cytoplasm at the animal pole. • A cap of cells called the blastodisc forms and begins gastrulation with the formation of the primitive streak. • In addition to the embryo, the three germ layers give rise to the four extraembryonic membranes: the yolk sac, amnion, chorion, and allantois. • The eggs of placental mammals are small and store little food, exhibiting holoblastic cleavage with no obvious polarity.
Gastrulation and organogenesis, however, resemble the processes in birds and reptiles. • After fertilization and early cleavage in the oviduct, the blastocyst implants in the uterus. • The trophoblast initiates formation of the fetal portion of the placenta, and the embryo proper develops from a single layer of cells, the epiblast, within the blastocyst. • Extraembryonic membranes homologous to those of birds and reptiles function in intrauterine development.
THE CELLULAR AND MOLECULAR BASIS OF MORPHOGENESIS AND DIFFERENTIATION IN ANIMALS
Morphogenesis in animals involves specific changes in cell shape, position, and adhesion • SEE Figure 47.16 and 47.17, p. 1012-1014 • Cytoskeletal rearrangements are responsible for changes in both shape and position of cells. • Both kinds of changes are involved in tissue invaginations, as occurs in gastrulation, for example. • The extracellular matrix provides anchorage for cells and also helps guide migrating cells toward their destinations. • Cell adhesion molecules on cell surfaces are also important for cell migration and for holding cells together in tissues.
The developmental fate of cells depends on cytoplasmic determinants and cell-cell induction.
Fate mapping can reveal cell genealogies in chordate embryos • SEE Figure 47.20, pp. 1014-1015 • Experimentally derived fate maps of embryos have shown that specific regions of the zygote or blastula develop into specific parts of older embryos.
The eggs of most vertebrates have cytoplasmic determinants that help establish the body axes and differences among cells of the early embryo • SEE FIGURE 47.21, p. 1015 • When cytoplasmic determinants are heterogeneously distributed in an egg, they serve as the basis for setting up differences among parts of the egg and, later, among the blastomeres resulting from cleavage of the zygote. • Cells that receive different cytoplasmic determinants acquire different fates.
Inductive signals drive differentiation and pattern formation in vertebrates • SEE FIGURES 47.22-47.24, pp. 1016-1019 • Cells in a developing embryo receive and interpret positional information that varies with location. • This information is often in the form of signal molecules secreted by cells in special "organizer" regions of the embryo, such as the dorsal lip of the blastopore in the amphibian gastrula and the apical ectodermal ridge of the vertebrate limb bud. • The signal molecules influence gene expression in the cells that receive them, leading to differentiation and the development of particular structures.