Download
1 / 1
20 likes | 174 Views
Mechanisms of Salinity Tolerance in Barley Z h onghua Chen 1 , Ian Newman 1 , Igor Pottosin 2 , Sergey Shabala 1 1 University of Tasmania and 2 Universi dad de Colima. Ian.Newman@utas.edu.au. After 24 h in 80 mM NaCl. 22 Na + influx, µ mol g -1 root FW. 80 mM NaCl at t=0.

E N D
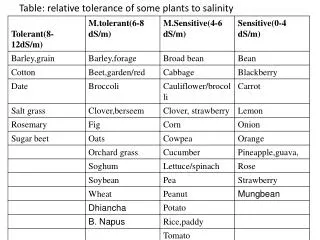
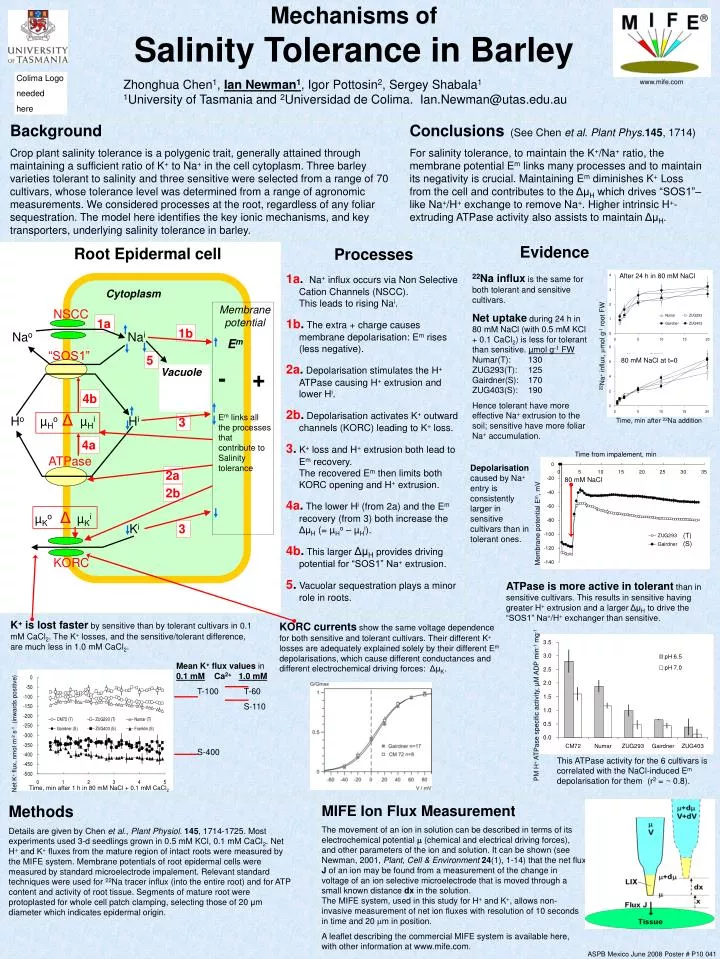
Mechanisms of Salinity Tolerance in Barley Zhonghua Chen1, Ian Newman1, Igor Pottosin2, Sergey Shabala11University of Tasmania and 2Universidadde Colima. Ian.Newman@utas.edu.au After 24 h in 80 mM NaCl 22Na+ influx, µmol g-1root FW 80 mM NaCl at t=0 Time, min after 22Na addition Time from impalement, min 80 mM NaCl Membrane potential Em, mV (T)(S) Mean K+ flux values in0.1 mM Ca2+1.0 mM T-100 T-60 S-110 S-400 Net K+ flux, nmol m-2 s-1 (inwards positive) PM H+ ATPase specific activity, µM ADP min-1 mg-1 Time, min after 1 h in 80 mM NaCl + 0.1 mM CaCl2 Colima Logo needed here www.mife.com Background Crop plant salinity tolerance is a polygenic trait, generally attained through maintaining a sufficient ratio of K+ to Na+ in the cell cytoplasm. Three barley varieties tolerant to salinity and three sensitive were selected from a range of 70 cultivars, whose tolerance level was determined from a range of agronomic measurements. We considered processes at the root, regardless of any foliar sequestration. The model here identifies the key ionic mechanisms, and key transporters, underlying salinity tolerance in barley. Conclusions (See Chen et al. Plant Phys.145, 1714) For salinity tolerance, to maintain the K+/Na+ ratio, the membrane potential Em links many processes and to maintain its negativity is crucial. Maintaining Em diminishes K+ Loss from the cell and contributes to the ΔμH which drives “SOS1”–like Na+/H+ exchange to remove Na+. Higher intrinsic H+-extruding ATPase activity also assists to maintain ΔμH. Evidence Root Epidermal cell Processes 1a.Na+ influx occurs via Non Selective Cation Channels (NSCC).This leads to rising Nai. 1b. The extra + charge causes membrane depolarisation: Em rises (less negative). 2a.Depolarisation stimulates the H+ ATPase causing H+ extrusion and lower Hi. 2b.Depolarisation activates K+ outward channels (KORC) leading to K+ loss. 3. K+ loss and H+ extrusion both lead to Em recovery. The recovered Em then limits both KORC opening and H+ extrusion. 4a. The lower Hi (from 2a) and the Em recovery (from 3) both increase the ΔμH (= µHo – µHi). 4b. This larger ΔμH provides driving potential for “SOS1” Na+ extrusion. 5. Vacuolar sequestration plays a minor role in roots. 22Na influx is the same for both tolerant and sensitive cultivars. Net uptake during 24 h in 80 mM NaCl (with 0.5 mM KCl + 0.1 CaCl2) is less for tolerant than sensitive. µmol g-1 FWNumar(T): 130ZUG293(T): 125Gairdner(S): 170ZUG403(S): 190 Hence tolerant have more effective Na+ extrusion to the soil; sensitive have more foliar Na+ accumulation. Cytoplasm Membranepotential Em - + Em links all the processes that contribute to Salinity tolerance NSCCNao Nai 1a 1b “SOS1” 5 Vacuole 4b HoµHo ΔµHi Hi 3 4a ATPase Depolarisation caused by Na+ entry is consistently larger in sensitive cultivars than in tolerant ones. 2a 2b µKo ΔµKi Ki 3 KORC ATPase is more active in tolerant than in sensitive cultivars. This results in sensitive having greater H+ extrusion and a larger ΔμH to drive the “SOS1” Na+/H+ exchanger than sensitive. K+ is lost faster by sensitive than by tolerant cultivars in 0.1 mM CaCl2. The K+ losses, and the sensitive/tolerant difference, are much less in 1.0 mM CaCl2. KORC currents show the same voltage dependence for both sensitive and tolerant cultivars. Their different K+ losses are adequately explained solely by their different Em depolarisations, which cause different conductances and different electrochemical driving forces: ΔμK. This ATPase activity for the 6 cultivars is correlated with the NaCl-induced Em depolarisation for them (r2 = ~ 0.8). Methods Details are given by Chen et al., Plant Physiol.145, 1714-1725. Most experiments used 3-d seedlings grown in 0.5 mM KCl, 0.1 mM CaCl2. Net H+ and K+ fluxes from the mature region of intact roots were measured by the MIFE system. Membrane potentials of root epidermal cells were measured by standard microelectrode impalement. Relevant standard techniques were used for 22Na tracer influx (into the entire root) and for ATP content and activity of root tissue. Segments of mature root were protoplasted for whole cell patch clamping, selecting those of 20 μm diameter which indicates epidermal origin. MIFE Ion Flux Measurement The movement of an ion in solution can be described in terms of its electrochemical potential m(chemical and electrical driving forces), and other parameters of the ion and solution. It can be shown (see Newman, 2001, Plant, Cell & Environment24(1), 1-14) that the net flux Jof an ion may be found from a measurement of the change in voltage of an ion selective microelectrode that is moved through a small known distancedx in the solution. The MIFE system, used in this study for H+ and K+, allows non-invasive measurement of net ion fluxes with resolution of 10 seconds in time and 20 m in position. A leaflet describing the commercial MIFE system is available here, with other information at www.mife.com. ASPB Mexico June 2008 Poster # P10 041