Download

1 / 23
430 likes | 1.07k Views
9 The Nucleus. Student Learning Outcomes: Nucleus distinguishes Eukaryotes from Prokaryotes Explain general structures of Nuclear Envelope, Nuclear Lamina, Nuclear Pore Complex Explain movement of proteins and RNA between Nucleus and Cytoplasm

E N D
9 The Nucleus • Student Learning Outcomes: • Nucleus distinguishes • Eukaryotes from Prokaryotes • Explain general structures of Nuclear Envelope, Nuclear Lamina, Nuclear Pore Complex • Explain movement of proteins and RNA between Nucleus and Cytoplasm Selective traffic of proteins, RNAs regulates gene expression [Describe the Internal Organization of the Nucleus] • Describe the Nucleolus and rRNA Processing
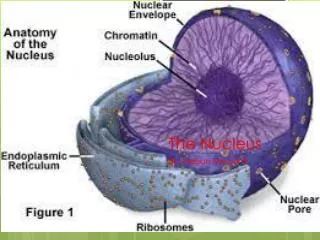
Nuclear Envelope;Traffic between Nucleus and Cytoplasm 1. Nuclear envelope: Two membranes Underlying nuclear lamina Nuclear pore complexes Outer membrane continuous with ER; membrane proteins bind cytoskeleton Inner membrane proteins bind nuclear lamina Fig. 9.1: EM of nucleus arrows indicate nuclear pores
Nuclear membrane, nuclear pores Fig. 9.1 Outer membrane is continuous with ER; Note ribosomes on ER Each nuclear membrane is phospholipid bilayer permeable only to small nonpolar molecules. Nuclear pore complexes are sole channels for small polar molecules, ions, proteins, RNA to pass through nuclear envelope. Fig. 9.2: EM of nucleus arrows indicate nuclear pores
Nuclear Envelope,Traffic between Nucleus and Cytoplasm Nuclear lamina is fibrous mesh (structural support): Fibrous proteins (lamins), andother proteins. Mutations in lamin genes cause inherited diseases Fig. 9.3: EM of nuclear lamina Hutchinson-Gilford Progeriacauses premature aging; Mutations in LMNA gene affect Lamin A protein
Nuclear Envelope, Traffic between Nucleus and Cytoplasm Mammals have 3 lamin genes, (A, B, and C), which code for at least 7 proteins. Two lamins form dimer, α-helical regions of 2 polypeptide chains wind around each other -> coiled coil. Lamin dimers associate to form nuclear lamina. Fig. 9.4
Nuclear Envelope, Traffic between Nucleus and Cytoplasm Nuclear pore complexes large 120 nm Complex: vertebrates, 30 different proteins (nucleoporins) Circular structures on faces of membrane; 8-fold symmetry. Lamina: loose mesh in nucleus Lamins bind: Protein emerin, lamin B receptor (LBR) (inner membrane) Chromatin. . Figs. 9.5, 9,7
Nuclear Pore,Traffic between Nucleus and Cytoplasm Nuclear pore complex - 8 spokes connected to rings at nuclear and cytoplasmic surfaces. Spoke-ring assembly surrounds central channel Protein filaments extend from rings: Basketlike structure on nuclear side. Cytoplasmic filaments on cytoplasmic side Fig. 9.8 nuclear Pore complex
Nuclear Pore Complex, Traffic between Nucleus and Cytoplasm Nuclear Pore Complex controls traffic between nucleus and cytoplasm: critical for physiology Passive transport: smallmolecules pass freely through channels Selective transport: energy-dependent for macromolecules (proteins and RNAs) Fig. 9.6 nuclear pore complex controls transport
Nuclear Envelope, Traffic between Nucleus and Cytoplasm Nuclear localization signals (NLS): Required for proteins to enter nucleus- specific aaseq Recognized by nuclear transport receptors transport of proteins through nuclear pore first identified on SV40 T antigen (viral replication protein) mutants helped figure Some NLS are one aaseq Others bipartitateseq A, kinase with SV40 NLS; B, mutated NLS
Nuclear Envelope, Traffic between Nucleus and Cytoplasm Import of proteins to nucleus: NLS recognized by nuclear transport receptors –importins Activity of nuclear transport receptors regulated by Ran, a GTP-binding protein Importins bind cargo at NLS sequence Move through pore Ran-GTP unloads, takes importin out. High concentration of Ran/GTP in nucleus: enzyme localization: GAP does GTP hydrolysis in cytoplasm GEF does GDP/ GTP exchange in nucleus (Fig. 9.20) Fig. 9.11 import of proteins
Nuclear Envelope, Traffic between Nucleus and Cytoplasm Nuclear export signals (NES): Required for proteins targeted for export Signals recognized by exportins (receptors in nucleus) direct transport to cytoplasm Less well characterized than NLS Ran also required for nuclear export Ran/GTP promotes binding of exportins and their cargo proteins, Ran/GTP dissociates complexes between importins and cargos (see Fig. 9.10) Fig. 9.12 export of proteins
Many importins and exportins are family of nuclear transport receptors - karyopherins.
Nuclear Envelope, Traffic between Nucleus and Cytoplasm Regulation of Protein transport is another point at which nuclear protein activity can be controlled: Regulation of import, export of transcription factors: Inhibitors block import (IkB and NF-kB) phosphorylation can block import (de-PO4 releases) Fig. 9.13 regulated import
Nuclear Envelope, Traffic between Nucleus and Cytoplasm Most RNAs are exported from nucleus to cytoplasm to function in protein synthesis: Active, energy-dependent process requires transport receptors Transported as ribonucleoprotein complexes (RNPs). rRNAs associate with ribosomal proteins, specific RNA processing proteins in nucleolus (Fig. 9.31). mRNAs associate with 20 proteins during processing, transport Fig. 9.14 EM of RNP transport : insect salivary gland; RNA unfolds
Fig 9.15 Transport of snRNAs between nucleus and cytoplasm Many small RNAs (snRNAs, snoRNAs) function in nucleus. • snRNAs are transported to cytoplasm by exportin (Crm1) • associate with proteins to form snRNPs and return to nucleus; snRNPs function in splicing Fig. 9.15 RNA
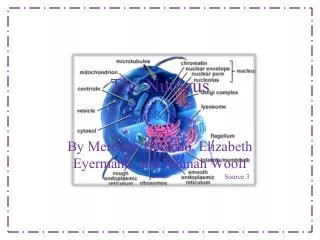
Internal Organization of the Nucleus 2. Internal structure of nucleus: organized, localized In animal cells, lamins where chromatin attachmes, organize other proteins into functional nuclear bodies Heterochromatin highly condensed, transcriptionally inactive; Euchromatin decondensed, all over Chromosomes organized in territories: Actively transcribed genes at periphery Fig. 9.16 arrow = nucleolus; arrowheads = heterochromatin Fig. 9.19 mammalian nucleus: DNA probes to chrom 4
Internal Organization of the Nucleus Nuclear processes appear localized (sequestered) to distinct subnuclear regions: DNA replication: Mammalian cells: clustered sites labeling newly synthesized DNA with bromodeoxyuridine (BrdU in place of T) Immunofluorescence (Ab to BrdU): newly replicated DNA in discrete clusters Fig. 21 – A: early replication B, late replication
Internal Organization of the Nucleus Nuclear processes appear localized (sequestered) to distinct subnuclear regions nuclear speckles: mRNA splicing machinery Detect with immunofluorescent staining - antibodies against snRNPs and splicing factors. PML bodies have transcription factors, chromatin-modifying proteins; identified from protein in promyelocytic leukemia Fig. 9.22 Speckles Fig. 9.23 PML bodies
The Nucleolus and rRNA Processing *3. Nucleolus:Site of rRNA transcription, processing, some aspects of ribosome assembly. Actively growing mammalian cells have 5 to 10 x 106 ribosomes, must be synthesized each time cell divides. Nucleolus is not surrounded by a membrane Multiple copies of rRNA genes (200 human) In oocytes, rRNA genes amplified, synthesis for early development. rRNA genes amplified 2000-fold in Xenopus oocytes, thousands of nucleoli, →1012 ribosomes per oocyte Fig. 9.26 XenopusoocyterRNA genes
The Nucleolus and rRNA Processing Nucleolar organizing regions: After each cell division, nucleoli reform, associated to genes for 5.8S, 18S, and 28S rRNA genes Each nucleolar organizing region has tandemly repeated rRNA genes separated by spacer DNA 5.8S, 18S, and 28S rRNAs are transcribed as single unit in nucleolus by RNA pol I → 45S ribosomal precursor RNA Fig. 9.28 Fig. 9.25
Fig 9.29 Processing of pre-rRNA Primary transcript of rRNA genes is large 45S pre-rRNA • pre-rRNA processed via series of cleavages, and some base modifications, including methylations • snoRNPs (snoRNAs with proteins) assemble on pre-rRNA as processing complexes (like spliceosomes on pre-mRNA) Fig. 9.29 ETS, external transcribed ITS, internal transcribed
Fig 9.31 Ribosome assembly • Formation of ribosomesrequires assembly of pre-rRNA with ribosomal proteins and 5S rRNA, then export of subunits • pol II made the mRNA for ribosomal proteins. Fig. 9.31
Review questions: 1. Eukaryote nuclear membranes separate transcription from translation. What regulatory mechanisms unique to eukaryotes achieve this regulation? 3. If you inject a frog egg with two globular proteins, one 15 kd and the other 100 kd, both of which lack NLS, will either protein enter the nucleus? 4. What determines the directionality of nuclear import? 5. Describe how the activity of a transcription factor can be regulated by nuclear import. * Consider the effect of mutations at gene level that inactivate NLS, NES, prevent phosphorylation of key sites, or prevent binding inhibitors on function