Download

1 / 55
550 likes | 654 Views
Role of Extracellular Matrix in Adaptation of Tendon and Skeletal Muscle to Mechanical Loading MICHAEL KJÆR Physiol Rev 84: 649–698, 2004; 10.1152

E N D
Role of Extracellular Matrix in Adaptation of Tendon and Skeletal Muscle to Mechanical Loading MICHAEL KJÆR Physiol Rev 84: 649–698, 2004; 10.1152 La matrice extracellulare (ECM) che si trova nei tessuti dei tendini, intorno e dentro ai muscoli, assicura il legame funzionale fra le cellule muscolari e l’osso. Segnali derivanti dal carico meccanico danno inizio ad una cascata di eventi che vanno dall’espressione genica alla transcrizione, translazione e processi post translazionali che infine regolano la sintesi proteica dell’ECM. L’ECM consiste di una varietà di sostanze, fra cui fibrille collagene e proteoglicani (PG) sono ubiquitari, ma ci sono anche PG non collageni. La forza di trasmissione del complesso muscolo-tendine dipende dall’integrità strutturale delle singole fibre muscolari e dell’ECM, dalla disposizione delle fibre nel tendine e quindi dalla sua capacità di assorbire e caricare energia. È inoltre noto che la resistenza alla tensione della matrice si basa su legami crociati intra- e intermolecolari, sull’orientamento, la densità e la lunghezza delle fibrille e delle fibre del collagene. Per meccanotrasduzione si intende un importante meccanismo in base al quale lo stress meccanico agisce sulle cellule scatenando segnali intracellulari, promuovendone la crescita e la sopravvivenza, regolandone la morfologia e l’architettura e influenzandone le risposte metaboliche.
È evidente che nei tendini e nei muscoli qualunque stimolo meccanico può dare inizio ad un adattamento che renderà il tessuto più resistente ai danni ottimizzando la trasmissione di forza. L’ECM è un conglomerato di sostanze le cui proprietà biochimiche e biofisiche costituiscono una rete flessibile che integra le informazioni sul carico convertendole in proprietà meccaniche. Funziona da supporto per l’adesione delle cellule mediata da integrine, distroglicano e proteoglicani sulla superficie cellulare e ai recettori della tirosinchinasi. L’interazione fra l’ECM e le molecole di adesione attiva segnali intracellulari e modifica il citoscheletro. L’ECM libera anche fattori di crescita. Un ruolo dominante nei rapporti funzionali fra l’ECM e le cellule è giocato dalle integrine, che costituiscono un vero e proprio organello sensibile allo stiramento. Le vie intracellulari della meccanotrasduzione compredono una chinasi di adesione focale (FAK), la paxillina, una chinasi legata all’integrina (ILK-1) e la proteinchinasi attivata dalla mitosi (MAPK). Quest’ultima è attivata sia dalla contrazione muscolare sia da forze applicate dall’esterno. I tendini sono un’organizzazione sistematica e fortemente compatta di tessuto connettivo dominata da collagene che è organizzato in fibrille, fibre, fasci di fibre e fascicoli, insieme ad altre componenti dell’ECM. Le cellule dei tendini sono fibroblasti, disposti in catene longitudinali con numerose appendici a fascia che si propagano nell’ECM e sono collegate attraverso gap junctions. Queste cellule in coltura rispondono agli stimoli meccanici producendo diversi componenti dell’ECM
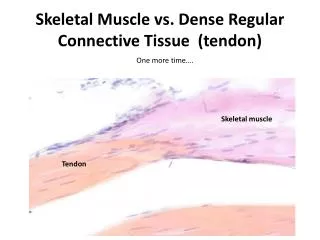
Sezione del tendine. Si vede un fascio di fibroblasti circondato da connettivo lasso. Immagine laser confocale: fibroblasti adiacenti in un fascicolo; le cellule hanno prolungamenti a fascia che le uniscono. Sezione trasversale e longitudinale con evidenziazione della proteina delle gap junctions connessina 43 (verde) e dei nuclei (rosso). Esiste una rete di comunicazione fra le cellule tendinee.
Rispetto ai muscoli, i tendini sono poco vascolarizzati: l’area occupata dai vasi non supera l’1-2%, ma il flusso di sangue durante l’esercizio può aumentare fino a 7 volte, che è comunque molto meno di quanto accade nel muscolo (20 volte). La regolazione del flusso nei tendini è indipendente da quella nei muscoli. Componenti ECM nel muscolo scheletrico Il tessuto connettivo intramuscolare ha molteplici funzioni: costituisce supporto meccanico a vasi e nervi; garantisce le risposte elastiche passive; la trasmissione di forza dalle fibre muscolari non si limita a tendini e ossa ma ha anche un andamento laterale fra cellule vicine e fra i fascicoli del muscolo. Il tessuto connettivo intramuscolare rappresenta un volume pari al 1-10% dell’intero muscolo, con molta variabilità fra muscoli. L’endomisio avvolge ogni fibra con fibre collagene sparse che permettono i movimenti durante la contrazione, il perimisio, fatto di fibre a spessore multiplo è trasversale rispetto alle fibre e le riunisce in gruppi, infine l’epimisio è composto di due strati di fibre collagene intrecciate e forma una guaina sulla superficie del tendine. Il connettivo intramuscolare è soprattutto collagene e serve non solo per organizzare le fibre in fascicoli ma anche alla trasmissione della forza. Vi sono diversi tipi di collagene: IV: sul sarco-lemma; I e III, in piccola misura V: epi- endo- perimisio; I: intramuscolare; gli altri tipi (II, VI, IX, XI-XVI e XVIII-XIX) sono presenti in minima quantità. Le diverse quantità relative dei vari tipi di connettivo in muscoli particolari sono associate alle funzioni e al ruolo del connettivo. Nel periodo perinatale la struttura del connettivo intramuscolare si modifica molto e l’organiz-zazione strutturale cambia in poche settimane, suggerendo un’interazione fra lo sviluppo del muscolo e del connettivo intramuscolare.
Struttura del connettivo intramuscolare: microscopia a scansione dopo rimozione delle proteine muscolari. In alto a destra ingrandimento dell’endomisio che circonda una singola fibra muscolare.
Ruolo funzionale dell’ECM nel tendine È importante riconoscere che il tendine e il connettivo intramuscolare interagiscono strettamen-te con gli elementi contrattili del muscolo per trasmettere la forza. Le dimensioni del tendine influenzano la loro capacità di essere stirati e di immagazzinare e rilasciare energia elastica durante i movimenti riducendo il costo energetico globale nel cammino e nella corsa. Diversi modelli di soppressione genica di singoli componenti dell’ECM nei topi riproducono vari aspetti della distrofia muscolare nell’uomo. Inoltre, l’imposizione di carichi compressivi modifica la composizione dell’ECM. REGOLAZIONE DEL COLLAGENE E DI ALTRE PROTEINE DELL’ECM: INFLUENZA DEL CARICO MECCANICO. La biosintesi del collagene è caratterizzata dalla presenza di un grande numero di modificazioni post traslazionali delle catene polipeptidiche, che determinano la qualità e la stabilità delle molecole. Nella cellula, la traslazione di mRNA del preprocollagene avviene nei ribosomi e l’assemblaggio del procollagene nel reticolo endoplasmatico. È noto che l’ECM del muscolo scheletrico risponde all’aumento di carico provocato dall’allena-mento di endurance, dall’esercizio acuto o da ipertrofia compensatoria sperimentale aumen-tando l’espressione e la sintesi di collagene nel muscolo. L’esercizio strenuo, in particolare il sollevamento di pesi che comprende componenti eccentriche provoca danno muscolare. L’esercizio acuto modifica l’attività metabolica e infiammatoria della regione peritendinea e aumenta la formazione di connettivo di tipo I durante il ricupero; quindi il carico acuto provoca adattamenti non limitati al tessuto osseo
Sintesi e degrado del collagene di tipo I. Elenco di potenziali fattori di crescita che regolano l’attivazione dei geni. TGF-b, transforming growth factor-.; IGF/IGF- BP, insulin-like growth factor and its binding proteins; IL, interleukin; FGF, fibroblast growth factor; PG, prostaglandin; VEGF, vasoactive endothelial growth factor. Mitogen-activated protein kinase (MAPK) ha un ruolo fondamentale nell’avviare l’attività genica mentre le metalloproteinasi (MMP) regolano la degradazione del collagene in funzione del carico meccanico
La risposta iniziale all’allenamento è un aumento del turnover del collagene I, seguito da più intensi processi anabolici, Lo schema di stimolazione della sintesi e del degrado con predominanza di processi anabolici è analogo a quanto accade alle proteine muscolari in risposta al carico. EFFETTI DELL’IMMOBILIZZAZIONE Nel ratto si è dimostrata una riduzione dell’attività enzimatica della biosintesi del collagene sia nel muscolo sia nel tendine, ma la quantità di collagene non cambia DEGRADAZIONE DEL TESSUTO CONNETTIVO NEL TENDINE E NEL MUSCOLO: EFFETTI DEL CARICO MECCANICO La degradazione del collagene è una tappa obbligatoria del turnover e del rimodellamento del tessuto in funzione del carico. Vi sono vie intra ed extracellulari che utilizzano rispettivamente la fagocitosi lisosomiale e le proteinasi dell’ECM. Nell’ambiente extracellulare la degradazione del collagene è effettuata da MMPs che sono generalmente presenti in forma di precursori, mentre questi enzimi (e i loro inibitori, le TIMPs) non contribuiscono alla fagocitosi lisosomiale. Le TIMPs sono spesso attivate contemporaneamente alle MMPs in risposta alla stimolazione fisica, indicando che c’è allo stesso tempo stimolazione ed inibizione delle degradazione. È probabile che le MMPs vengano prima e che le TIMPs funzionino come regolatori che limitano la degradazione. La possibilità di allungare i tendini varia dall’1-3% (m. estensore) al 3-4% (m. flessore), fino al 16% del tendine d’Achille (coniglio). In cadaveri umani la massima estensibilità arriva al 6%. La parte libera del tendine permette accumulo e rilascio di energia elastica mentre l’aponeurosi serve alla trasmissione vera e propria delle forze contrattili.
Il rimodellamento dei tendini comprende aspetti quantitativi (sezione) e/o qualitativi (resistenza alla tensione). Gli effetti dell’allenamento sono variabili sulle due componenti che spesso non coesistono Con l’invecchiamento e nel diabete i legami crociati non specifici (mediati dalla condensazione di uno zucchero riducente con gruppi aminici) aumentano la glicazione delle proteine tendinee portando ad una maggiore resistenza tensile ma ad un irrigidimento. Tutto questo riduce la capacità di adattamento e il turnover del collagene e stimola i fibroblasti aumentando la fibrosi. È dimostrato che lo stiramento passivo non modifica le proprietà viscoelastiche dei tendini. Gli adattamenti indotti dal carico sull’ECM, in particolare la sintesi di collagene, dipendono da ormoni regolatori e fattori di crescita e anche da alcuni canali ionici. Ruolo fondamentale di FGF, TGF-b, IL-1, IL-6, IGF, IGF-BP (binding-protein)
An overview of the endocrinology of skeletal muscle Melinda Sheffield-Moore and Randall J. Urban TRENDS in Endocrinology and Metabolism Vol.15 No.3 April 2004 Gli ormoni sono fondamentali regolatori del metabolismo muscolare in condizioni normali e patologiche. Il muscolo scheletrico dell’uomo arriva al 40-45% del peso corporeo ed è uno dei tessuti più attivi metabolicamente, ma le influenze metaboliche che da esso derivano sono spesso trascurate: il muscolo è una grossa riserva di substrati metabolici ed un importante bersaglio degli ormoni. Il muscolo è un deposito praticamente infinito di proteine e aminoacidi e fornisce precursori del glucosio attraverso neoglucogenesi. L’accumulo di proteine si verifica solo se la sintesi supera la degradazione, mentre c’è perdita quando avviene il contrario. Il bilancio omeostatico del muscolo dipende da: alimentazione, esercizio, età, malattia. Il processo di rimodellamento è influenzato dagli ormoni: testosterone, GH, insulina, IGF-I e glucocorticoidi. Ormoni anabolizzanti: stimolano la crescita del muscolo aumentando la sintesi delle proteine, riducendone la degradazione o facendo entrambe le cose. Gli androgeni hanno diverse azioni biologiche perché hanno come bersaglio sia tessuti riproduttivi, sia altri tessuti, come i muscoli. Malattie che aumentano il catabolismo muscolare (AIDS) correlano con gli androgeni circolanti. Si stima che il 50% dei maschi >50 anni siano ipogonadici, anche se spesso asintomatici. L’uso di agenti anabolizzanti per l’ipogonadismo tipico o per quello traumatico è clinicamente giustificabile.
Lo scopo principale della terapia ormonale negli anziani è di mantenere o migliorare la forza muscolare. Il testosterone in anziani ipogonadici rallenta la degradazione proteica e aumenta la sintesi, ma spesso aumenta molto di più la massa muscolare che la forza: questa tende comunque a diminuire di più della massa negli anziani. Anche le donne subiscono queste alterazioni muscolari con l’età: i livelli di testosterone circolante diminuiscono di circa la metà da 20 a 40 anni. La terapia sostitutiva è efficace nelle donne pre menopausa, ma molto di più dopo 65 anni. GH e IGF-1 – nonostante credenze popolari sulle straordinarie proprietà del GH i suoi effetti sull’anabolismo sono modesti. L’abuso ne deve comunque essere scoraggiato, ma i suoi effetti potenziali vanno ancora studiati. Supplementazioni di rhGH per 5 anni hanno aumentato la forza dei flessori del ginocchio e la presa della mano, con maggiori effetti sui muscoli prossimali e minori nelle femmine: data la lentezza della risposta, rispetto a quella degli androgeni, l’effetto anabolizzante del GH è stato indicato come minimo. Numerose controindicazioni derivano dall’effetto neoglucogenetico, che induce al diabete. Più importante la sua azione stimolante sull’IGF. Insulina e muscolo scheletrico – L’insulina non si limita a regolare il metabolismo glucidico e lipidico, ma interferisce con il metabolismo proteico nel muscolo, per il quale è un potente stimolo, purché siano disponibili aminoacidi.
Estrogeni e muscolo scheletrico – Gli effetti degli estrogeni sono stati poco studiati. Benché il muscolo possieda recettori per gli estrogeni, questi non hanno sostanzialmente effetti sulla massa e forza muscolare nelle donne in menopausa. Gli estrogeni anche di origine extragonadica, anche nei maschi, sono molto importanti per l’omeostasi dell’osso. Ormoni catabolici – glucocorticoidi e muscolo scheletrico. Insulti traumatici gravi, in particolare le ustioni, iniziano una risposta ipermetabolica, con forte aumento del cortisolo e diminuzione di androgeni: parte la degradazione proteica, con fortissimo turnover e fuoruscita di aminoacidi dalle riserve intracellulari. La perdita di massa magra non si contrasta nemmeno con una dieta iperproteica. Miostatina e muscolo scheletrico – La miostatina (GrowthDifferentationFactor-8 ) appartiene ai TGF; è una proteina che limita l’accrescimento dei muscoli, dove può raggiungere elevate concentrazioni: la sua importanza è stata riconosciuta quando si è capito che la soppressione del suo gene caratterizza una razza di bovini che hanno il doppio della massa muscolare; topi con deficienza di miostatina hanno muscoli ipertrofici ed inoltre hanno maggiore atrofia da immobilizzazione. La somministrazione di cortisolo stimola la produzione di miostatina.
Functional, structural and molecular plasticity of mammalian skeletal muscle in responsetoexercisestimuli Martin Flück The Journal of Experimental Biology 209, 2239-2248 La malleabilità del muscolo scheletrico che ne permette il rimodellamento strutturale secondo le esigenze è un fenomeno particolarmente impressionante nel regno animale. Questa plasticità si traduce negli adattamenti di forza, resistenza e velocità in seguito a variazioni dei carichi: questo accade negli atleti nei quali le modificazioni fenotipiche che massimizzano le prestazioni specifiche sono impressionanti. Un aspetto importante della plasticità è la sua specificità per cui l’intensità del carico e il numero delle contrazioni muscolari costituiscono gli stimoli per gli adattamenti. Quindi il profilo delle perturbazioni muscolari esercita un controllo essenziale sul fenotipo muscolare. Questa rassegna indica le scoperte più recenti sul coinvolgimento dell’espressione genica in risposta a stimoli di esercizio ripetitivi. Adattamento all’allenamento di endurance – Le fibre si differenziano verso un fenotipo con alto volume mitocondriale, aumentata densità capillare e deviazione verso fibre lente. Nell’insieme, queste modificazioni contribuiscono alla massimizzazione del trasporto di substrati, all’aumento della capacità respiratoria e delle caratteristiche contrattili nel corso di contrazioni lente e frequenti che caratterizzano l’esercizio di endurance. Alla base degli aggiustamenti muscolari sta l’espressione genica in risposta all’attività fisica: singole perturbazioni omeostatiche indotte dall’esercizio si integrano nella produzione di copie di geni diffusibili (mRNA) che stimolano la traslazione, nell’apparato ribosomico, delle proteine codificate.
Integrazione di stimoli fisiologici nella risposta fenotipica. Le perturbazioni omeostatiche indotte dall’esercizio si integrano regolando le alterazioni della trascrizione genica. Le copie di geni diffusibili istruiscono il rimodellamento muscolare mediante l’assemblaggio delle proteine codificate.
Processi metabolici nelle fibre muscolari. Il processo biochimico principale che genera energia nel muscolo è la combustione di acidi grassi e carboidrati. I carboidrati sono impor-tati attraverso processi facilitati dai capillari alle miocellule dove sono depositati come trigliceridi o glicogeno intracellulare per esse-re utilizzati in seguito. La metabolizzazione degli acidi grassi è un processo obbligato-riamente aerobico che avviene nei mitocondri attraverso beta ossidazione e ciclo di Krebs. Al contrario, la conversione metabolica dei carboidrati per glicolisi nel citoplasma è indipenden-te dall’ossigeno e non è necessariamente legata alla respirazione dei mitocondri. Questo porta alla produzione anaerobica di lattato. La scomposizione delle catene organiche nei mitocondri produce equivalenti riducenti (e CO2), che portano alla generazione di ATP ossigeno-dipendente, accoppiata alla catena respiratoria. Le sigle nei riquadri sono proteine cruciali per il trasporto, il deposito e la conversione nelle vie metaboliche del muscolo. LPL endoteliale entra nel trasporto di acidi grassi (FA) dai vasi attraverso l’interstizio fino al compartimento intracellulare. H-FABP trasporta FA nella cellula. HSL libera FA da IMCL. CPTI è un enzima indispensabile per l’assun-zione di FA nella matrice mitocondriale. Gli enzimi del ciclo di KrebsFum e SDH e i costituenti della catena di trasporto degli elettroni NADH6, COX1 e COX4 realizzano la produzione di ATP. PFKM rappresenta un passaggio principale per l’ingresso dei carboidrati nella via glicolitica.
I livelli di mRNAdelle principali subunità respiratorie mitocondriali sono significativamente correlati con il volume dei mitocondri, quindi l’adattamento locale del numero dei trascritti mitocondriali è co-regolato e accoppiato alla respirazione massimale del sistema (VO2max) durante l’esercizio. Analisi del tibiale anteriore di biatleti competitivi e controlli rivela aumento contemporaneo di transcritti genici dei fattori per la mobilizzazione degli acidi grassi. Quindi l’allenamento aerobico migliora la respirazione mitocondriale e aumenta la capacità del muscolo di importare acidi grassi dal sangue. La combinazione degli adattamenti espressionali in tappe successive della via ossidativa nei muscoli allenati riflette il livello locale di coordinamento che contribuisce al miglioramento sistemico delle capacità aerobiche. La corrispondenza delle variazioni di espressione genica con adattamenti funzionali della via ossidativa dopo allenamento indica un preciso circuito molecolare nel muscolo che sta alla base dell’aumentata resistenza alla fatica. Questa relazione implica uno specifico meccanismo che coordina gli adattamenti genici in base al reclutamento dei muscoli. L’adattamento del tessuto muscolare all’aumento dell’attività contrattile avviene probabilmente nella fase di ricupero dopo ogni episodio di esercizio defatigante. Questo comporta un eccesso di adattamenti cellulari che produce l’accumulo di risposte rimodellanti incrementali dopo ogni episodio e ripetendo gli esercizi si arriva al miglioramento della performance. Con tecniche particolari si può seguire l’andamento nel tempo della produzione di mRNA: si è dimostrata una transitoria stimolazione genica 8 ore dopo un esercizio al cicloergometro di 30 min. Al contrario, una volta che gli adattamenti muscolari siano stabili dopo 6 settimane di allenamento questa risposta genica è attenuata: questo è dovuto all’aumento stabile dei livelli di mRNA e spiega la progressiva riduzione dell’efficacia dell’allenamento in soggetti allenati.
Microadattamenti dell’espressione di trascritti in relazione all’allenamento. Modello dell’aumento dell’RNA e della performance con la ripetizione dell’esercizio. Ogni episodio provoca un eccesso di livello di trascrizione nelle fasi di ricupero. Le linee rappresentano l’evoluzione dei livelli di RNA e della performance
Timecourse della risposta muscolare all’esercizio in condizioni di normo (A) e ipossia (B). In A e B sono riportati le variazioni significative di trascritti della via ossidativa. I principali fattori del citoscheletro, titina e desmina non sono modificati, a dimostrare la specificità ella risposta
La stimolazione della trascrizione degli enzimi legati al metabolismo dei grassi è attenuata nell’esercizio in ipossia. Gli aggiustamenti strutturali e funzionali (rimodellamento) durante l’allenamento derivano dall’accumulo di risposte nell’espressione genica agli esercizi ripetuti. Il complesso stimolo costituito dall’esercizio provoca una serie di perturbazioni omeostatiche nei muscoli interessati. Esse sono recepite come precisi segnali di processo e si traducono nell’attivazione della trascrizione genica o nella sua stabilizzazione. Le perturbazioni sono di tipo metabolico, meccanico e ormonale. Sintetizzando i risultati presentati si conclude che una complessa risposta dei geni riflette la specificità dell’adattamento muscolare a diversi tipi di esercizio. Vi è una co-regolazione della via ossidativa, dei cromosomi e dei genomi. L’evidente correlazione fra famiglie di geni e le relazioni struttura-funzione rivela l’esistenza di un preciso circuito molecolare che regola la via del metabolismo ossidativo e di un meccanismo a soglia (switch = interruttore) che coordina le risposte locali all’allenamento
Listofabbreviations ACADL long-chainacyl-CoAdehydrogenase ACADVL very-long-chainacyl-CoAdehydrogenase ACADM medium-chainspecificacyl-CoAdehydrogenase ALDOA aldolase A ALDOC aldolase C AMPK 5.AMP-activated proteinkinase CPT1 carnitine palmitolytransferase 1 CYC cytochrome c COX1 cytochrome c oxidasesubunit 1 COX4 cytochrome c oxidasesubunit 4 COX5B cytochrome c oxidasesubunit 5B ECH1 enoyl-CoAhydratase FFA free fattyacids Fumfumarase GAPDH glyceraldehyde-3-phosphate dehydrogenase G-6-P glucose-6-phosphate HADH 3-hydroxyacyl-CoA dehydrogenasetype II HADHB 3-hydroxyacyl-CoA dehydrogenase B H-FABP fatty acid binding protein of the heart HIF-1.,. hypoxia-induciblefactor 1.,. HSL hormone-sensitivelipase IMCL intramyocellularlipids PFK phosphofructokinase JNK c-jun N-terminal kinase LDH3 lactate dehydrogenase 3 LDL R low-density lipoprotein receptor LPL alkaline lipoprotein lipase NADH nicotinamide adenine dinucleotide dehydrogenase PFKM phosphofructo-kinase muscle-type PFKFB1 6-phosphofructo-2-kinase/fructose-2,6- biphosphatase 1 PPAR. peroxisome proliferator-activated receptor . PPAR. peroxisome proliferator-activated receptor . SDH succinate dehydrogenase Tfam mitochondrial transcription factor PGC-1. peroxisome proliferator coactivator-1. VLDLR Very low-density lipoprotein receptor WT wild type
http://www.unm.edu/~lkravitz/index.html The Mystery of Skeletal Muscle HypertrophyRichard Joshua Hernandez, B.S. and Len Kravitz, Ph.D. L’esercizio, lavoro muscolare compiuto contro un carico progressivamente più grande, provoca l’aumento della massa e della sezione dei muscoli: questo si chiama ipertrofia. Ma perché la cellula muscolare cresce e come fa a crescere? Nonostante una mole notevole di ricerche, non è ancora del tuttochiaro il quadro completo (e molto complesso) del graduale adattamento del muscolo a carichi crescenti. L’ipertrofia muscolare è l’aumento della massa e dell’area di sezione dei muscoli. L’aumento di dimensioni è dovuto ad un aumento dello spessore (non della lunghezza) di tutte le fibre del muscolo. Sia il muscolo scheletrico sia quello cardiaco si adattano a carichi di lavoro regolari crescenti che superano le capacità pre-esistenti del muscolo. Il cuore aumenta la sua funzione di pompa mentre il muscolo scheletrico diventa più efficiente nel trasmettere forza ai segmenti ossei attraverso i tendini. Le cellule satellite facilitano l’accrescimento, la manutenzione e la riparazione del muscolo danneggiato (non il cuore). Sono chiamate satelliti perché si trovano sulla superficie esterna delle fibre muscolari fra il sarcolemma e la lamina basale; sono mononucleate e quasi prive di citoplasma. Normalmente sono dormienti e sono attivate da qualsiasi tipo di trauma o danno muscolare . Quando si svegliano proliferano e si spostano sul muscolo danneggiato, si fondono con le fibre preesistenti aumentando il materiale nucleare che serve a rigenerare la fibra.
Il numero di cellule satelliti presenti in un muscolo dipende al tipo di fibre muscolari: le fibre lente ne hanno 5 o 6 volte di più di quelle veloci. Il trauma provocato da esercizi di resistenza (soprattutto eccentrici) mette in moto il sistema immunitario: invasione di macrofagi con secrezione di citochine, fattori di crescita e altre sostanze, che richiamano altre cellule del sistema immunitario. Le tre citochine importanti in questo contesto sono: IL-1, IL-6, TNF (chiamate citochine proinfiammatorie). Promuovono la dissociazione delle proteine, la rimozione di detriti cellulari e la produzione di prostaglandine. I fattori di crescita sono proteine altamente specifiche che stimolano la differenziazione e la divisione di determinate cellule. Particolarmente importanti: IGF, FGF e HGF (epatociti). Questi fattori collaborano nella stimolazione delle cellule satelliti. Ormone della crescita (GH). Stimola l’IGF e l’attivazione, proliferazione e differenziazione delle cellule satelliti. Gli effetti ipertrofici dell’assunzione di GH come anabolizzante sembrano legati più a ritenzione di liquidi che all’aumento delle proteine contrattili. Cortisolo. Ormone steroideo legato allo stress che stimola la gluconeogenesi anche attraverso il catabolismo proteico. Per quel che riguarda l’ipertrofia, inibendo la sintesi delle proteine esso ha un effetto negativo.
Testosterone. È il principale ormone androgeno e ha i maggiori effetti sugli organi sessuali maschili, ma anche su sistema nervoso, muscolo scheletrico, midollo osseo, cute e peli del corpo. Sui muscoli ha un importante effetto anabolico, che contribuisce anche alle differenze di genere sulla composizione corporea. Stimola l’anabolismo proteico e pertanto l’ipertrofia. Fibre di tipo I. Si ipertrofizzano notevolmente con sovraccarichi progressivi, non solo con l’esercizio di resistenza ma anche in qualche misura con l’esercizio aerobico. Fibre di tipo IIa. Le loro percentuale aumenta con l’allenamento per trasformazione di IIb in IIa. Il disuso provoca la trasformazione inversa. Fibre di tipo IIb. Si convertono in IIa con l’esercizio di resistenza che aumenta complessivamente le capacità ossidative del muscolo migliorando la resistenza alla fatica.
La prima cellula satellite di mammifero: sezione traversa di una fibra del sartorio di ratto … si vedono le membrane contigue di una cellula satellite (sp) e di una fibra muscolare (mp); la membrana basale (bm) si prolunga sopra la congiunzione delle membrane delle due cellule (microscopio elettronico x22.000)
Known knowns, known unknowns and unknown unknowns Conclusioni: fatti noti, consapevolmente non noti e inconsapevolmente non noti Cosa sappiamo dunque dopo 50 anni di ricerca sulle cellule satellite del muscolo? Sostanzialmente, che si tratta di cellule staminali residenti, che forniscono mioblasti per l’accrescimento del muscolo, l’omeostasi, l’ipertrofia e la riparazione. Poco si sa di come siano mantenute in uno stato quiescente, mentre si comincia a capire come siano attivate per entrare nel ciclo cellulare. Una modalità relativamente nuova è la regolazione dei geni da parte di miRNA. Analogamente, si cominciano ad avere informazioni sul ruolo della nicchia sublaminaledelle cellule satelliti. È molto controverso se le cellule satelliti siano eterogenee o costituiscano un unico pool, perché alcune si differenziano mentre altre si moltiplicano conservando la riserva di cellule staminali.
Skeletalmuscleremodeling Matthew J. Potthoff, Eric N. Olson and RhondaBassel-DubyCurrent Opinion in Rheumatology 2007, 19:542–549 Il muscolo scheletrico sembra omogeneo nei preparati istologici, ma invece si compone di una popolazione eterogenea di miofibreche differiscono per proprietà metaboliche e contrattili. Le miofibre sono classificate sulla base dell’espressione genica delle catene pesanti della miosina (MHC). Le fibre di tipo I, lente, esprimono MHC I/b, si contraggono lentamente sono ricche di mitocondri e miosina, hanno metabolismo ossidativo e sono resistenti alla fatica. Le fibre di tipo II, veloci, esprimono MHC di tipo II, si affaticano rapidamente, si contraggono velocemente e hanno metabolismo glicolitico o ossidativo. L’espressione genica delle miofibre può cambiare in risposta alle richieste dell’ambiente mediante l’attivazione di segnali trasduzionali che promuovono variazioni adattative della citoarchitettura e della composizione proteica. Le miofibre primarie che esprimono miosine di tipo adulto o embrionale si stabilizzano precocemente durante lo sviluppo, mentre le miofibre adulte si stabilizzano dopo la nascita e le miosine adulte sostituiscono quelle embrionali. Hanno un alto grado di plasticità e possono rimodellarsi fenotipicamente in risposta a stimoli specifici, come il carico contrattile, l’ambiente ormonale e malattie sistemiche. Per esempio, l’allenamento trasforma preesistenti fibre veloci nel fenotipo ossidativo e al contrario una riduzione dell’attività neuromuscolare provoca la trasformazione inversa.È stato stabilito che specifici schemi di impulsi condotti dai nervi motori esercitano un’influenza fenotipica sui muscoli che innervano
La concentrazione intracellulare di calcio si eleva in risposta a stimoli fisiologici e attiva il fattore nucleare calcineurina della cellule T attivate (NFAT) e la via fattore stimolante i miociti 2 (MEF2)-istone deacetilasi (HDAC), PGC-1, peroxisome proliferator-activated receptor coactivator-1; RCAN1, regulator of calcineurin 1.
L’attivazione della via IGF/Akt/mTOR(mammalian target of rapamycin) stimola l’ipertrofia e antagonizza la perdita di massa muscolare. Nell’uomo Akt, mTOR e glicogeno sintetasi aumentano in seguito ad un allenamento di resistenza che induce ipertrofia muscolare. mTOR è una chinasi attivata da Akt e regola la sintesi proteica sulla base della disponibilità di sostanze nutritizie. È un elemento importante nella regolazione della massa muscolare dopo la nascita specialmente quando aumenta il carico sui muscoli. L’atrofia muscolare che capita con l’invecchiamento, l’immobilizzazione, i voli spaziali, i traumi muscoloscheletrici e molte malattie cataboliche, è dovuta all’aumento della distruzione proteica mediato dalla via ubuquitina proteasoma. Sono aumentati due geni: muscle RING finger 1 (MuRF1) and muscle atrophy F-box (MAFbx/atrogin-1). Vi sono molte prove che uno stato infiammatorio prolungato (età e malattie croniche) provoca perdita di muscolo. Questo indica che le terapie che controllano l’infiammazione possono anche curare l’atrofia muscolare.
THE MOLECULAR BASIS OF TRAINING ADAPTATION (Coffey VG & Hawley JA, Sports Med 2007; 37: 737) CONTROL OF THE SIZE OF THE HUMAN MUSCLE (Rennie MJ, Wackerhage H, Spangerburg EE & Booth FW, Annu Rev Physiol 2004; 66: 799) I muscoli che non lavorano perdono massa e forza (ipotrofia, atrofia; sarcopenia) I muscoli sottoposti a tensione (attiva – contrazione, passiva – stiramento) aumentano massa e forza (ipertrofia) I muscoli sottoposti ad attività prolungata di bassa intensità aumentano la resistenza alla fatica e il consumo di ossigeno
ALLENAMENTO (TRAINING) Effetti specifici dell’aumento di una funzione specifica: miglioramento della funzione specifica • VARIABILI: • intensità • durata • frequenza
Distruzione/ rimozione Distruzione/ rimozione Ricostruzione/sintesi proteica Ricostruzione/sintesi proteica segnali segnali Dimensioni/funzioni EFFETTI: da carico in eccesso (overload) Tutte le cellule si modificano (remodeling)
ALLENAMENTO DELLA FORZA Effetto della prima contrazione massimale o ripetizioni submax Forte dipendenza da condizioni iniziali: scarsa attività detraining Fattore principale: volume muscolare, sezione trasversa
ALLENAMENTO DELLA FORZA Importanza degli elementi non contrattili Importanza della disposizione del muscolo es. pennazione La forza aumenta anche per migliore attivazione delle unità motorie
ALLENAMENTO DELLA FORZA La massima forza si esprime con una contrazione eccentrica Per contrazioni isometriche 1/2 forza Nelle contrazioni concentriche la forza è inversamente proporzionale alla velocità Un movimento reale non può essere perfettamente isotonico, perché la rotazione intorno alle articolazioni modifica le leve
ALLENAMENTO DELLA FORZA Effetti: • Sulla massa muscolare: • ipertrofia • iperplasia • Sul fenotipo (tipi di fibre) • Sul controllo motorio
ALLENAMENTO DELLA FORZA Ipertrofia • Aumento dell’area di sezione: effetto diretto sulla forza sviluppata • Aumento di lunghezza, per aggiunta di sarcomeri in serie: spostamento sulla curva tensione/lunghezza
ALLENAMENTO DELLA FORZA Iperplasia • Aumento del numero di fibre: dimostrato solo in alcuni modelli animali • Ruolo delle cellule satellite: vengono sicuramente attivate dallo sforzo • Più importanti per l’ipertrofia: si fondono con le fibre mature, fornendo materiale strutturale, in particolare nuclei (e mitocondri?)
ALLENAMENTO DELLA FORZA Ruolo del SNC Strategie: • Aumento della frequenza di scarica: tetano completo. • Treni d’impulsi: frequenza impulsi nel treno • durata treno • aumento frequenza treni • stimolazione tetanica continua • Reclutamento di unità motorie
Distruzione/ rimozione Ricostruzione/sintesi proteica Forza Resistenza () segnali ALLENAMENTO DELLA FORZA Biologia molecolare Tournover proteico: 1 g/kg/dì; semivita 7-15 gg
ALLENAMENTO AEROBICO MECCANISMI CENTRALI Adattamenti cardiaci > volume plasmatico > concentrazione Hb (emoglobina) *** anemia relativa dell’atleta: spiegazione aumento 2,3 DPG inganno meccanismo eritropoietina Coronarie: densità capillare; flusso (ruolo NO) Migliore distensibilità ventricolare in diastole
ALLENAMENTO AEROBICO MECCANISMI periferici > densità capillare (30-40%): migliore diffusione gas; ridotta velocità di transito Mitocondri: > numero > enzimi ossidativi capacità ossidativa > uso FFA: ruolo di LPL endoteliali migliore economia energetica (ossidativa) Accumulo (relativo) di glicogeno
ALLENAMENTO AEROBICO MECCANISMI periferici Aumento estrazione ossigeno (Da-v O2) Aumenta flusso muscolare massimo (iperemia metabolica): ruolo NO Aumenta concentrazione mioglobina
Stem cells in postnatal myogenesis: molecular mechanisms of satellite cell quiescence, activation and replenishment J. Dhawan and T.A. Rando TRENDS in cell biology (2005) 15:666-673 Cellula satellite: principale cellula staminale nel m. scheletrico. Fra la lamina basale e il sarcolemma Stato di quiescenza: non si differenziano, non si moltiplicano Attivazione fortissima per: esercizio, danno muscolare, malattie degenerative
Attiva ricostituzione delle cellule satelliti dopo attivazione per poter ripetere processi rigenerativi Lo stato di quiescenza non è una condizione di semplice inattività, ma di controllo dei processi trascrizionali Espressione di proteine tipiche (riconoscimento) – Miostatina: mantiene la condizione di quiescenza Alterazioni della nicchia che contiene le cellule satelliti ne provocano l’attivazione: stadio intermedio altamente proliferativo (cell. Progenitrici)
Nei soggetti anziani le cellule satelliti sono meno facilmente attivabili, ma quando attivate proliferano normalmente: il rallentamento correlato con l’età è legato a fattori modificabili della nicchia cellulare Il numero di cellule satelliti rimane costante nell’adulto: alla prima moltiplicazione cellulare, solo una delle cellule figlie continua la differenziazione, mentre l’altra rimane allo stadio staminale
Mechanotransduction and the regulation of protein synthesis in skeletal muscle T.A. Hornberger and K.A. Esser Proc. Nutr. Soc. (2004) 63: 331-335 La tensione regola la massa muscolare, anche in vitro: aumenta la sintesi proteica e diminuisce specularmente la degradazione Capacità intrinseca del muscolo scheletrico, a prescindere da fattori di regolazione come: testosterone, glucocorticoidi, fattori di crescita
Meccanocezione: accoppiamento di informazione meccanica con eventi biochimici intracellulari. Interessa: A) la membrana; B) il complesso matrice extracellulare-integrina-citoscheletro A) la membrana: bistrato lipidico allo stato semiliquido; resiste alla deformazione. Un aumento della superficie del 2-4% rompe la membrana. La membrana è sovrabbondante (100%), formando pieghe, microvilli, caveole. Questa è una riserva che previene la rottura