Download

1 / 56
560 likes | 570 Views
Learn the journey of sound from mechanical vibrations in the ear to electrical signals in the brain. Explore the anatomy, physiology, and function of the auditory system, including the outer, middle, and inner ear structures.

E N D
Introduction to the Auditory System Joseph Santos-Sacchi
Acoustical mechanical electrical Outer/middle/inner inner/neural
Sound is a disturbance that propagates through air, or any medium. It is absent in a vacuum. There is a compression and rarefaction of the molecules that comprise the medium. The speed of sound is about 350 m/s in air. The magnitude (degree of compression) and frequency (reciprocal of period) of acoustic waves that travel through space can be represented by a sinusoid. All sound, no matter how complex, is composed of component sinusoids. We hear from about 200-20,000 hertz (cycles per second), corresponding to low through high pitch. The magnitudes scale used for hearing is logarithmic, and is relative to the faintest sound we can hear. The ratio reanges up to about 120 decibels, above which the perception becomes painful. Normal conversational speech is about 60 decibels.
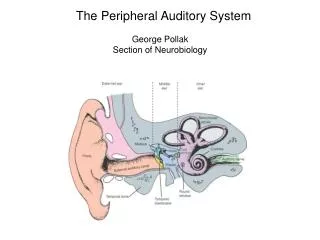
This classic drawing by Brodel illustrates the three components of the auditory periphery. The outer ear, middle ear and inner ear. The pinna and ear canal (outer ear) funnels sound to the tympanic membrane (ear drum), which vibrates and causes the middle ear ossicles (bones) to vibrate in turn. The malleus contacts the tympanic membrane, and transmits the vibrations to the incus, and stapes. The stapes footplate inserts into the oval window of the fluid filled cochlea. This arrangement ultimately causes vibrations within the fluids of the cochlea.
Without the middle ear, sound would effectively impinge directly on the fluids of the inner ear. 99.9% of the sound energy would be reflected. It is the job of the middle ear to get that energy to the cochlear where the hair cells can respond. Two processes account for the middle ear’s transformer activity; they are the increased pressure at the oval window afforded by a ratio advantage between tympanic membrane area and stapes footplate area, and the lever ratio of the arm lengths of the incus and malleus. The area ratio concept can be appreciated by considering how easily the pointy part of a fat nail can be pushed into a piece of wood. The combined effect is significant – with middle ear disease or disarticulated bones hearing losses up to 40 decibels can be obtained.
The picture on the left shows the membranous cochlea after the temporal bone was chipped away. Blue arrow is oval window where stapes footplate inserts. White arrow is the round window. The right picture is a scanning electron micrograph of the cochlea showing the organ of Corti’s spiral path from base to apex.
Upper left shows the cochlea coil sliced open. The coil is actually composed of three spiraling tubes, The innermost tube, the scala media, houses the sensory epithelium and other important structures.
The organ of Corti rests on the basilar membrane and has a variety of cell types; there are two basic classes – hair cells and supporting cells. Supporting cells metabolically and physically support the hair cells. There is one row of inner hair cells and 3-4 rows of outer hair cells. Total number is about 12-14 thousand hair cells, which do not regenerate when destroyed. The high K (140 mM) and positive potential (+80 mV) found in the scala media (SM) are important for normal hair cell transduction, since it provides the drive that governs K+ flow through the stereociliar transduction channels. Remember there is a voltage gradient of about 150 mV across the stereociliar of outer hair cells (OHCs) (intracellular resting potential of –70 mV vs, +80 mV in SM) and a gradient of about 125 mV across the stereociliar of IHCs (intracellular resting potential of –45 mV vs, +80 mV in SM).
Actual section through the guinea pig organ of Corti. The hair cells are identified.
A quick ultrastructural tour of important cochlear cellular elements
So let’s take an anatomical tour of the cochlea. We can see the stapes footplate (S) inserting in the cochlea’s oval window. The bone normally overlying the cochlea is chipped away to see the spiraling tubes.
Still higher magnification. Looking down on the top of the organ of Corti, we see the stereocilia bundles that protrude from the 3 rows of OHCs and one row of IHCs. P, top of pillar cells; D, top od Deiters; cells.
Looking down on the organ of Corti, but the organ is cracked open so we can see the bodies of the OHCs. Remember this is the hair cell type that dances wildly when it hear salsa music! Imagine all of them going nuts in your ears. Under the OHCs are Deiters (D) cells that support the OHCs. Process of a Deiteres cell(P).
Very high magnification of the top of an OHC. The stereocilia bundle (S) is composed of numerous stereocilia that protrude from the hair cell. They range in height. The arrangement allows the hair cell to respond selectively during movements of the bundle towards and away from the tallest stereocilia.
Top of longitudinal section of single stereocilium. Actin filaments are packed within the plasma membrane boundaries.
Base of stereocilia, where their rootlets penetrate into the actin rich cuticular plate at the apical pole of the hair cell.
Stereocilia are stiff. Here a glass coverslip pushes them down from above. They keel over like little telephone poles.
When the bundle is moved toward the tallest stereocilia the hair cell depolarizes from its normal resting potential and the spike activity of the innervating afferent fiber increases. The opposite occurs when the bundle is moved in the opposite direction.
This depicts the response of the cell (the intracellular receptor potential) when the bundle is deflected in different directions. Only when it is deflected towards and away from the tallest stereocilia is the response maximal (0 and 180 degrees). If the deflection is a right angles (90 degrees) there is no response.
If the bundle is wiggled back and forth with a sinusoidal stimulus, then the receptor potential is sinusoidal as well. The response due to displacement is sigmoidal, with excitatory bundle movements producing positive voltage responses, and inhibitory movements producing negative voltage responses. However, the response magnitude for a given displacement in either direction is not symmetrical. Because of this, the receptor potential will have a sinusoidal component (ac) and an offset component in the depolarizing direction (dc). Remember a depolarization of the hair cell is excitatory.
Stereocilia have ionic channels that open and close when mechanically stimulated. The tip links (red arrows in schematics and electron micrograph) connect from one stereocilium to another; when the two stereocilia shear past each other during bundle displacement, the link pulls on an ionic channel gate which opens and allows ions (mainly K+) to flow inside the cell, causing depolarization. Since there are no chemical reactions, the gating process is very fast and can follow sound-induced perturbations tens of thousands of times per second.
In the organ of Corti bundle movements are caused when the basilar membrane vibrates up and down due to a sound stimulus. This results from relative shear movements between tectorial membrane (TM, where stereocilia insert) and the tops of the hair cells, causing the bundles to wiggle
Normally the stereocilia are stiff rods, but after loud noise exposure they may become temporarily damaged and floppy. Hearing loss and ringing in the ear (tinnitus) can occur. If the sound exposure was not too severe, full recovery is possible.
How does sound cause the hair cells to be stimulated? When the stapes vibrates in the oval window, it caused the fluids to be displaced, which in turn causes a deflection of the basilar membrane.
The basilar membrane (BM; here uncoiled) is not uniform; its stiffness varies along the length of the cochlear spiral, with the base region (near the stapes) stiffer than the apex. Remember that stiff structures tend to vibrate at higher frequencies than less stiff structures.
Due to the mechanical characteristics of the BM, different frequencies of sound cause a maximal vibration at different positions along the BM length. High frequencies cause a vibration that is maximal near the base, and low frequencies cause a maximal vibration near the apex. The basilar membrane performs a mechanical frequency analysis – this is the basis of tonotopicity – the mapping of frequency responsiveness along an anatomic structure. When a sound containing two or more frequencies (e.g. speech) stimulates the basilar membrane each frequency component will cause a maximal vibration at its characteristic place.
Since hair cells rest on the basilar membrane, hair cells at a particular location will be “tuned” – respond maximally – to a particular frequency. The graph on the left shows the response from an inner (white squares) and outer (black squares) hair cell when stimulated with different frequencies of the same magnitude. Each cell produces a maximal response near 1 kilohertz (kHz).
Hair cells, in this case an inner hair cell, are innervated by the eighth cranial nerve – the auditory nerve. The synapse is a chemical one, and uses a transmitter similar to glutamate. Depolarization cause the release of neurotransmitter into the synaptic cleft, and upon binding to receptors on the postsynaptic membrane cause the afferent fiber to depolarize. If threshold is reached, the fiber will fire an action potential.
Inner hair cells receive the majority of afferent fibers from the eighth nerve, whereas outer hair cells only have about 5%.
How might the auditory system encode pitch and loudness? Frequency – pitch Intensity – loudness
So, if the hair cell is tuned to a particular frequency then it is obvious that the nerve fiber that innervates it will also be tuned similarly. Here we plot the response threshold (the smallest intensity of sound required to cause an increase in spike rate above the spontaneous activity) from a bunch of eighth nerve fibers that arise from different regions of the basilar membrane. Each fiber responds bets to a particular frequency range, the more limited that range the better the tuning. We see that tuning is better for the high frequencies than for the low frequencies.
Here we plot the tuning curve for a nerve fiber (based on spike threshold, dotted line) and the basilar membrane (based on mechanical vibration, solid lines), and we see that they are both sharply tuned. This illustrates that it it the tuning of the basilar membrane that underlies frequency selectivity throughout the auditory system – from hair cell on up.
Outer hair cell isolated from the guinea pig organ of Corti. It is about 10 um wide. At the top are the stereocilia; near the bottom the round nucleus can be seen. This slide introduces the player in the following video. In the original video, the OHC was stimulated via an intracellular electrode with the electrical signal from Tito Puente’s salsa music. Note that the music was not played acoustically, but the electrical signal directly stimulated the lateral membrane of the cell. In the following digital recording, the cell responds to Linda Ronstadt’s music.
Click the image to play. You may have to raise you volume to hear the music!
The first synapse of the eighth nerve (AN) occurs in the cochlea nucleus (red circle, CN).
There are several types of cells in the cochlea nucleus, but we will only focus on one – the bushy cell. This cell type has a very large and efficient synapse which allows the cells to faithfully reproduce and transmit the activity of the eighth nerve.
The response from the cells types of the cochlear nucleus are varied and depend on the membrane characteristics of the cells as well as on local interaction via interneurons. Note that the bushy cell types that have large synapses have a response pattern that is primary-like.
The superior olivary complex is composed of the lateral superior olive (LSO), the medial superior olive (MSO) and the medial nucleus of the trapezoid body (MNTB). It is the first location where sound information from both ears is integrated.
The binaural properties gained at the level of the SOC, while certainly relayed centrally, are not simply propagated. Interactions at higher levels occur, e.g., between nuclei of the lateral lemniscus of either side. In general, characteristics first observed in the lower brainstem are enhanced further up. A sharpening of tuning, e.g. to time differences, is found between SOC and IC. Binaural localization is not lost when SOC is ablated.
Auditory radiations project from the MGB of the thalamus to the auditory cortex. The primary areas are located in Brodman’s areas 41 and 42. Three regions comprise the primary areas, and receive direct input from the thalamus. There are other cortical association areas that receive input from the primary areas. You will hear about Wernicke’s area (speech reception) and Broca’s area (speech production) during the language lecture. Strokes in these areas devastate communication abilities.
Note that although the anatomical layout differs in the auditory pathway, tonotopicity which was established at the level of the basilar membrane extends all the way to cortical regions.
Cortical columns (or sheets) in the primary auditory areas responds similarly. That is, an electrode traversing the cell layers of the cortex will respond similarly to a given characteristic, whether it be any of the four listed above.