Download
1 / 28
280 likes | 475 Views
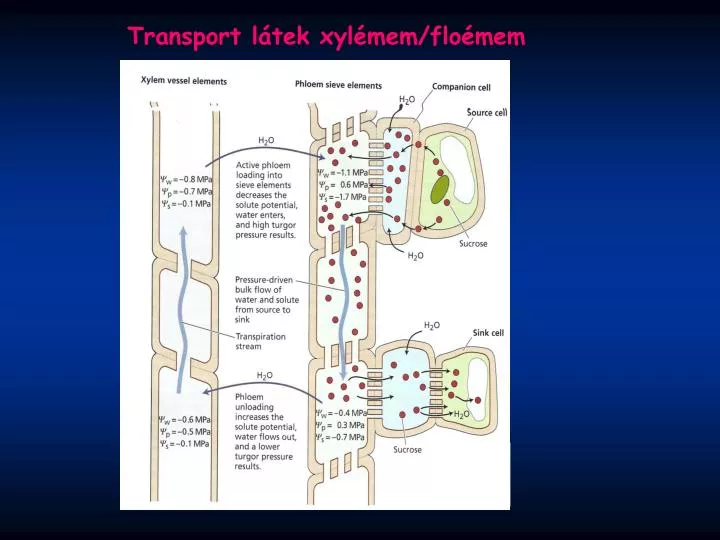
Transport látek xylémem/floémem. Transportéry. Transportéry a jejich role u „ mix/apoplastic loaders/unloaders “. Sacharózové transportéry (SUC, SUT (např. AtSUC1, HvSUT4 ).

E N D
Transportéry Transportéry a jejich role u „mix/apoplastic loaders/unloaders “ Sacharózové transportéry (SUC, SUT (např. AtSUC1, HvSUT4 ) • Nakládánído floému, vykládánído určitých sinků (svěrací buňky, vyvíjející se pylová zrna, vyvíjející se embrya /semena, na sacharidy bohaté plody… ) Expreseve zdrojových listech, většina v sincích, exprese některých genů vázaná pouze na určité pletivo (např.NtSUT3 jen v pylu a pylových láčkách) Účastní se také výměny látek při symbióze (mykorhiza, Rhizobium), ale i při napadení patogenem. Směr transportu sacharózy může být pravděpodobně měněn a závisí na sacharózovém gradientu, pH a transmembránovém potenciálu. Definitivní důkaz účasti transportéru při nakládání do floému: SUT1 u bramboru v antisence orientaci (také u Arabidopsis a tabáku) Kühnetal., 2003 Blokování transportu z listů zvýšení konc. sacharózy a škrobu v listech - omezení růstu, chlorotické léze, menší semena, časná senescence
4 jedno a dvouděložné 1 dvouděložné dvouděložné 5 jednoděložné 3 jednoděložné 2 jednoděložné jedno a dvouděložné jedno- i dvouděložné Fylogenetický strom sacharózových transportérů Sauer, 2007, Payyavula et al., 2011 Kühn a Grof, 2010 Transport sacharózy- kotransport s H+
4 jedno a dvouděložné 1 dvouděložné 5 jednoděložné 3 jednoděložné 2 jedno a dvouděložné Fylogenetický strom sacharózových transportérů Sauer, 2007 Kühn a Grof, 2010 Transport sacharózy- kotransport s H+
Malé rodiny SUT:(Arabidopsis 9genů – dva pseudogeny,, 7 transportéry, podle podobnosti spadají do tří subrodin, rýže 5 genů ) Subrodina 1:dvouděložné - vysoká afinita, - exprese v CC nebo SE buňkách (AtSUC1, AtSUC2, AtSUC5, AtSUC8, AtSUC9,) Subrodina 3:jednoděložné - vysoká afinita, exprese v CC nebo SE buňkách ( AtSUT2) prodloužená centrální smyčka, lokalizace do CC a SE zdroje ve vegetativních sincích Subrodina 2 jednoděložné dvouděložné nižší afinita AtSUT2 senzor ? Subrodina 4: ( AtSUT4) Vakuoly, plasmalema ? AtSUC 2 AtSUC4 Subrodina 5: jen necharakterizované transportéry jednoděložných Některé SUT nejsou specifické pro transport sacharózy, transportují také např. maltózu, turanózu, některé glukosidy, nebo i syntetické deriváty jako sukralóza – chlorovaná sacharóza, ne rafinózu Všechny dosud charakterizované … H+ sympotréry se stechiometrií 1:1 • rodina transportérů SWEET Usnadněná difúze, 7 transmembránových domén.. (A. thalina 17 genů) Otázka ???Jak je transportována sacharóza do vakuoly ??? Kühn a Grof, 2010; Schneider et al., 2012; Doidy et al., 2012; Doidy et al., 2012
Transportéry Regulace SUT/SUC Sacharózový transport přísně regulován • Regulace s vývojem listu : mladý list 0 exprese, vývoj listu exprese (vodivá pletiva), začíná v listových špičkách – pokračuje k bázi. Neúplná vazba na vývoj vodivého pletiva – zastínění - 0 exprese. • Regulace exprese pod kontrolou přísunu asimilátů z MC • Aktivita nepřímo úměrná turgorovému tlaku v SE Za sucha -- pokles tlaku… aktivity transportérů… stabilizace tlakového gradientu Při nedostatečném přísunu asimilátů pokles tlaku kompenzován akumulací iontů • regulace vnějšími podmínkami – teplotou, světelnými podmínkami, fotoperiodou, stresem, napadením patogenem..
Mechanismy regulace SUT/SUC • regulace exprese • regulace na posttranskripční úrovni- regulace pomocí miRNA (u všech At SUT s výjimkou At SUT2 a AtSUT4) Regulační role StSUT4 .. (ovlivnění kvetení a tuberizace) • fosforylace proteinu (AtSuc 5; AtSUC1) • redox-regulace – cílení a dimerizace při oxidativním prostředí- zvýšení aktivity (StSUT1, LeSUT1) • regulace interakcí protein-protein (interakce s proteiny v membráně, ale i v ER) • Předpokládá se regulace fytohormony (ABA), ale mechanizmus ???
Transport StSUT1mRNA/SUT1 do SE Předpoklad : transport SUT do SE je předmětem regulace SUT1 mRNA několika druhů nalezena ve floémové tekutině ??? Mobilní signál ?? Kühn and Grof, 2010
Př.: Regulace kvetení a tuberizace u bramboru StSUT4 působí - • Přes StSUT1 na export sacharózy z listů • Přes COL3 na syntézu mobilního signálu FT • Přes ovlivnění syntézy giberelinů Chincinska et al., 2008
Transportéry Monosacharidové transportéry Arabidopsis Hexózové transportéry Dvě genové rodiny: • rodina monosacharidových (MST) transportérů Regulace cytosolické koncentrace redukujících cukrů Hexózové transportéry se většinou přímo transportu na dlouhou vzdálenost neúčastní ALE!!! Nepřímo jej výrazně ovlivňují Arabidopsis : MST transportéry- genová rodina o 53 členech ! Thompson, 2006 Slewinski, 2011
Fylogenetický strom monosacharidové transportéry • 53 genů Arabidopsis (černá) • genů Medicagotrunculata (modrá) • 68 genů rýže (červená) GSL =ERD Doidy et al., 2012
Transportéry Hexózové transportéry MST lze rozdělit do 7skupin (uvnitř skupiny podobnost 42-96%), 12 transmembránových domén Skupina 1. Nejlépe prostudovaná skupina (značeníSTP-sugar transportprotein ) – Arabidopsis 14 genů hexózové transportéry plazmatické membrány, H+ symportéry, transport hexóz z apoplastického prostoru do buněk AtSTP1, vysoká afinita, transportuje monosacharidy, ne fruktózu lokalizace: klíčící semena; kořeny semenáčků; svěrací buňky děloh, listy růžice, stonky, kališní lístky exprese: stoupá za tmy, denní rytmus s vrcholem kolem poledne :H+ symportéry Vysoká afinita (kromě AtSTP3) Široké spektrum monosacharidů (kroměAtSTP9 a AtSTP14) Role ve vývoji sinků (kromě AtSTP3) AtSTP 2 lokalizace: pyl, AtSTP 6lokalizace: pyl AtSTP 9lokalizace: pyl (jediný specifický pro glukózu) AtSTP 3: nízká afinita lokalizace: zelená pletiva, poranění, útok patogena AtSTP 11: lokalizace: pyl AtSTP 4 : vysoká afinita lokalizace: špičky kořenů, pylové láčky, listy, exprese stoupá po poranění a napadení patogenem AtSTP 13: lokalizace: vodivá pletiva korunních lístků, indukce během PCD AtSTP 14: specificky transportuje galaktózu Transportované cukry: glukóza, fruktóza, galaktóza, manóza, xylóza
Transportéry Skupina 2:VGT(Vacuolar glucose transporters)- Arabidopsis 3 geny, transportuje glukózu AtVGT 1 a 2 na tonoplastu antiport s H+ AtVGT 3 ? chloroplast Skupina 3: TMT– (tonoplast monosaccharidetransporters)– Arabidopsis3 geny, tonoplastové glukózové transportéry, transport za stresových podmínek Skupina 4:pGlcT– plastidové glukózové transportéry Arabidopsis4 geny Možná spražený transport s H+, u jednoho prokázána lokalizace na Golgi, (Přes vysokou identitu- různé funkce) Skupina 5: PMT(Polyol/monosacharide transporters) Arabidopsis6 genů Transportují širokou škálu polyolů a monosacharidů, sorbitol, manitol, xylitol, xylóza, glukóza, fruktóza, galaktóza Skupina 6:INT (Inositol transporters) Arabidopsis 4 geny vysoce specifické myo-inositol/H+ symportery na membráně, na tonoplastu, exprese v pylu a v CC , pravděpodobně ? syntéza RFO Skupina 7:ERD-like – (early-responsive to dehydratation)- pasivní ale selektivní, glukóza, fruktóza, galaktóza, manóza, xylóza, nejméně znalostí, mobilizace výtok z vakuoly Arabidopsis19 genů SWEET-malé proteiny (Arabidopsis -17 genů), 7 trasnsmembránových domén, oboustanný uniport glukózy, lokalizace na plazmatické membráně, Slewinski, 2011
ERD Doidy et al., 2012
Transportéry Cukr v xylému Hexózové transportéry nalezeny v xylému V zimě - floémový tok téměř zastaven, rostliny v dormanci sacharidy v xylému, prevence proti zmrznutí a poškození Na jaře – obnova funkce (embolismus) – transport mobilizovaných zásob v kmeni a kořenech Javor cukrový
Transportéry • ???? Otázky kolem hexózových transportérů • Jak se děje transport do vakuoly ??? • Proč tolik různých transportérů v pylu? • Proč v pylu hexózové transportéry ? • Jak fruktóza z rozštěpené sacharózy vstupuje do buňky ? • Jak se liší funkce u těch lokalizovaných na stejné místo ? • Regulace jak na transkripční, tak post-transktipční úrovni ? • Malý efekt vyřazení jednotlivých transportérů ??? • Jak je zajištěn transport glukózy přes plastidové membrány ??? • Jak jsou vykládány RFO .. účast hexózových transportérů ???
Transportéry Př.:Transgenní rostliny bramboru Transport z listu (nakládání do floému) apoplastem nebo symplastem ?
Transportéry Př.:Transgenní rostliny bramboru Transport z listu (nakládání do floému) apoplastem nebo symplastem ? transformace : exprese invertázy v buněčné stěně ve zdrojovém listu MC floém sacharóza invertáza hexózy Redukovaný růst kořenů Redukovaný výnos hlíz Hromadění cukrů a škrobu v listech Transport do floému za účasti apoplastu, transport sacharózy
Transportéry Př.:Transgenní rostliny bramboru Transport z listu (nakládání do floému) apoplastem nebo symplastem ? Transport asimilátů (vykládání floému) do buněk sinku ?? transformace : exprese invertázy v buněčné stěně ve zdrojovém listu MC floém sacharóza invertáza hexózy Redukovaný růst kořenů Redukovaný výnos hlíz Hromadění cukrů a škrobu v listech Transport do floému za účasti apoplastu, transport sacharózy
Transportéry Př.:Transgenní rostliny bramboru Transport z listu (nakládání do floému) apoplastem nebo symplastem ? Transport asimilátů (vykládání floému) do buněk sinku ?? transformace : exprese invertázy v buněčné stěně ve zdrojovém listu transformace : exprese invertázy v buněčné stěně v sinku- hlíze MC floém floém parenchym sacharóza sacharóza invertáza invertáza hexózy hexózy Redukovaný růst kořenů Redukovaný výnos hlíz Hromadění cukrů a škrobu v listech Zvýšená akumulace hexóz Zvýšení výnosu hlíz Transport do floému za účasti apoplastu, transport sacharózy Transport do sinku – účast apoplastu, transport hexóz inhibice sacharózového transportéru v sinku mírný výnosu Př. : kukuřice – mutant bez invertázové aktivity v sinku maličká semena
Protonové pumpy: H+ ATPázy plazmatické membrány • Podmiňují sekundární aktivní transport přes membrány • Účastní se kontroly získávání a rozdělování živin po rostlině Arabidopsis thaliana (AHA) Otázka ? Jedna buňka může mít i několik isoforem genů ? Proč ? AHA 1– všude, více v nadzemních částech AHA 2– všude, více v kořenech AHA 3 – všude, zejména v CC AHA 4 – všude, hlavně v endodermis v kořenech AHA 5 - nízká exprese všude, snad specif. pro svěrací. b. Svěrací buňky :Vicia faba, tabák: 2 isoformy Arabidopsis – všech 11, nejvíc AHA 1,2,5 AHA 6 - prašníky AHA 7 – výhradně pyl Změny aktivity v závislosti na vnějších faktorech: Aktivace: salinita, chladová aklimace Inhibice: houbové elicitory, nízká teplota, vyčerpání cukrů AHA 8 – výhradně pyl AHA 9 - prašníky AHA 10 – nejvíce v šešulích AHA 11 – všude Regulace exprese na úrovni transkripce/ translace: velmi malá !!! AHA 12 – pseudogen, pylu Hlavní regulace : posttranslační změny !
Transport látek floémem Posttranslační změny • Fosforylace/defosforylace • Fosforylace C-koncové domény Vazba 14-3-3 proteinu aktivace N-doména vážící nukleotid A - řídící doména P- fosforylační doména Padersenetal., 2007
Transport látek floémem Posttranslační změny • Fosforylace/defosforylace • Fosforylace C-koncové domény Vazba 14-3-3 proteinu aktivace Hexamer: ATPáza + 14-3-3 protein Ottmann et al., 2007
Transport látek floémem Otázka ? Proč regulace na úrovní posttranslační ?? Posttranslační změny • Fosforylace/defosforylace • Fosforylace C-koncové domény Vazba 14-3-3 proteinu aktivace • Modulační fosforylace Hexamer: ATPáza + 14-3-3 protein • Inhibiční fosforylace Ottmannetal., 2007 • Změna množství enzymu Otázka ? Jaké kinázy a fosfatázy? Jaká jejich regulace ?? • Změna lokalizace v membráně
Zdrojová buňka periferní V1 –8 subjednotek (A,B,C,D,E,F,G,H) integrální V0- 6subjednotek a,c,c´c´´, d,e) Nedostatečné informace o tonoplastových transportérech, exprese identifikovaných hlavně v sincích, Transport do vakuoly Aktivní transport do vakuoly … závislý na protonovém gradientu (transport antiportem s H+) důležitá role H+ pump Vakuolární H+-ATPázy („starší“ a komplikovanější ) V1 - hydrolýza ATP V0 - translokace protonu. dva komplexy Subjednotky – s výjimkou subjednotek „VHA-a“ a „WHA-c“ jsou ostatní subjednotky často kódovány jedním genem Ztrátové mutace některých genů se projeví významně, jiné se téměř neprojeví Lokalizace také na Golgi Interace subjednotky A s 14-3-3 proteinem závislá na fosforylaci AHA-B1 nalezena v jaderném komplexu s HXK ! Účastní se glukózové signalizace
Zdrojová buňka Vakuolární H+-PPázy (H+ pyrofosfatázy) Jedna subjednotka, funkční homodimer ??? Jaký je fyziologický význam těchto rozdílů v citlivosti?? lokalizace: vakuola, Golgi, plazmatická membrána Dva typy – typ I – závislá na K+ v cytosolu, mírně citlivá k Ca2+ inhibici typ II – necitlivá k K+, extrémně citlivá k Ca2+ inhibici Oba typy vyžadují Mg2+ jako kofaktor Nalezen interakční motiv pro vazbu s 14-3-3 proteinem
Zdrojová buňka A B Vakuolární H+-PPázy (H+ pyrofosfatázy) Jedna subjednotka, funkční homodimer ??? Jaký je fyziologický význam těchto rozdílů v citlivosti?? lokalizace: vakuola, Golgi, plazmatická membrána Dva typy – typ I – závislá na K+ v cytosolu, mírně citlivá k Ca2+ inhibici typ II – necitlivá k K+, extrémně citlivá k Ca2+ inhibici Oba typy vyžadují Mg2+ jako kofaktor Nalezen interakční motiv pro vazbu s 14 -3-3 proteinem, ??? Jaké další funkce?? ?? Ovlivňuje přesun PM H+-ATPáz na plazmatickou membránu. ?? Vysoká citlivost k IAA ?? Přenos IAA signálu ??? ?? Aktivace ostatních H+ pump ?? Př:Rajče s expresí H+ PPázy typu I z Arabidopsis: Odolnost k nedostatku vody, patrně díky většímu rozvoji kořenového systému, podobné výsledky i u jiných rostlin. Př: Arabidopsis, H+ PPáza typu I, A – nadexprese B – ztrátová mutace ! U jiných ekotypů výrazně menší odezva