Download
1 / 31
310 likes | 479 Views
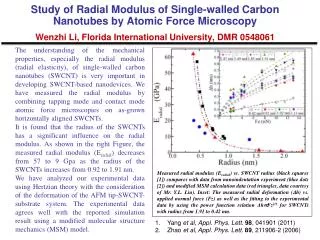
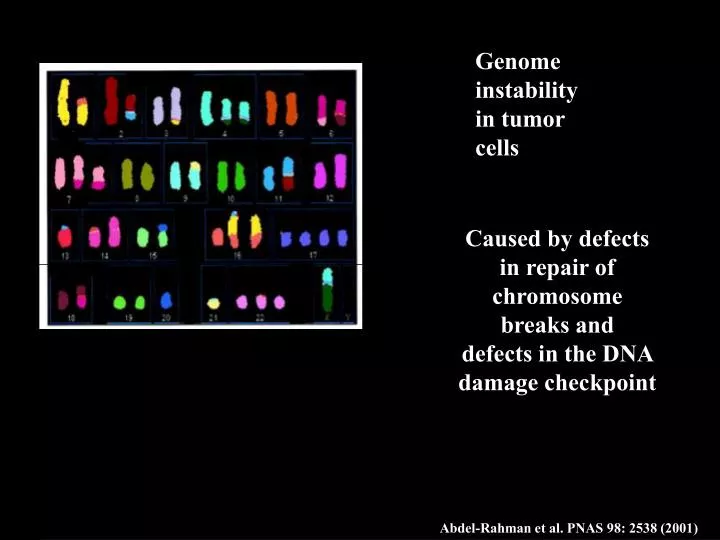
Caused by defects in repair of chromosome breaks and defects in the DNA damage checkpoint. Genome instability in tumor cells Truncations Translocations Inversions Duplications Amplifications Deletions Mutations. Abdel-Rahman et al. PNAS 98: 2538 (2001).

E N D
Caused by defects in repair of chromosome breaks and defects in the DNA damage checkpoint Genome instability in tumor cells Truncations Translocations Inversions Duplications Amplifications Deletions Mutations Abdel-Rahman et al. PNAS 98: 2538 (2001)
In vertebrates, repair of chromosome breaks is essential for life. Double-strand breaks (DSBs) arise during normal DNA replication. DSBs arising in chicken DT40 cells after Rad51 is depleted (Takeda lab)
DSB HO, I-SceI
How are double-strand breaks repaired? In vivo biochemistry what happens to the DNA? what proteins are doing what?
Saccharomyces MAT switching: a model for DSB-induced recombination and repair HO endonuclease
Saccharomyces MAT switching: a model for DSB-induced recombination and repair Galactose-induced HO endonuclease
0 1 2 4hr Physical monitoring of gene conversion by Southern blot analysis MATa MATa HO cut Connolly, White and Haber 1988
200 kb away HMLa MATa 5’ to 3’ resection (MRX and ?) Strand invasion (RPA, Rad51 etc) Primer extension (PCNA and Pold and e) 3’ nonhomology removal (Ercc1-XPF) Strand displacement (Srs2) Second strand synthesis All the newly synthesized DNA in the recipient locus HMLa MATa
Rad51p 0 0.3 0.5 1 2 3 5 hr Y Y a HO cut Visualizing recruitment of Rad51 to MAT by chromatin immunoprecipitation Neal Sugawara, Xuan Wang
Rad51p 0 0 0.3 0.3 0.5 0.5 1 1 2 2 3 3 5 hr 5 hr Y Y a HO cut Visualizing recruitment of Rad51 to MAT by chromatin immunoprecipitation Crosslink proteins to DNA with formaldehyde; shear chromatin; immunoprecipitate with anti-Rad51 antibody; reverse crosslinks; PCR amplify regions of interest MAT ChIP HML IP Neal Sugawara, Xuan Wang
Rad51p 0 0 0.3 0.3 0.5 0.5 1 1 2 2 3 3 5 hr 5 hr Y Y a HO cut Visualizing recruitment of Rad51 to MAT by chromatin immunoprecipitation Crosslink proteins to DNA with formaldehyde; shear chromatin; immunoprecipitate with anti-Rad51 antibody; reverse crosslinks; PCR amplify regions of interest MAT ChIP HML IP 10’ delay Neal Sugawara, Xuan Wang
Rad51p RPA 0 0 0.3 0.3 0.5 0.5 1 1 2 2 3 3 5 hr 5 hr Y Y a HO cut Visualizing recruitment of Rad51 and RPA to MAT by chromatin immunoprecipitation Rad51 at MAT HML IP RPA at MAT Neal Sugawara, Xuan Wang
0 0 0.3 0.3 0.5 0.5 1 1 2 2 3 3 5 hr 5 hr distal Y Y a HO cut Visualizing synapsis between MAT and HML by chromatin immunoprecipitation of Rad51 Non MAT DNA MAT IP HML IP Xuan Wang, Neal Sugawara
MATa Watching synapsis between MAT and HML Debra Bressan
This correlates well with results using chromatin immunoprecipitation for Rad51 recombinase 0 20’ 30’ 1 2 3 5 hr MAT HML 60 50 Cells with a single GFP spot (%) 40 30 20 10 0 0 60 120 180 240 300 360 Time after HO induction (min) Suvi Jain Debra Bressan
How does the cell know which donor to chose? donor preference
Donor Preference is controlled by the Recombination Enhancer that acts over 50 kb to control “accessiblity” of the left arm of chromosome III. RE binds both forkhead and SCF transcription factors, but there are no transcripts nearby! RE Donor preference during MAT switching 85-90% MATa HMLa HMRa MATa HMLa HMRa
MATa or MATa re∆ MATa RE HML MAT HML MAT HMR HMR HMRa RE CEN MATa We believe Fkh1 and Sw14/Swi6 make contact with “tethering sites” to change the mobility or position of the left arm of chromosome III
Biophysics of live chromosome movement! In collaboration with Jané Kondev (Physics) Opportunities for biophysics students! Susannah Gordon-Messer
The DNA damage checkpoint Preventing cell cycle progression to allow more time for DNA repair
HO ( ) ( ) Analysis of homologous recombination (HO “on” for 1 hr) HML MAT When HO endonuclease is continuously on HO 0.2% > 99% cells have a single broken chromosome
arrest adaptation HO endonuclease unrepaired DSB 1 DSB recovery DSB repaired
arrest HO endonuclease 1 DSB recovery How is the checkpoint turned off when DNA is repaired?(Recovery) DSB repaired
Checkpoint-mediated megabase-wide phosphorylation of histone H2AX in mammalian cells after DNA damage What about yeast?
THR4 60' Distribution of -H2AX in the region around the DSB What limits the spreading? Are there “barriers”? 25 MAT 60' 20 relative ChIP 15 10 5 0 170 180 190 200 210 220 230 240 250 chromosome location (kb) Jung-Ae Kim
Kinetics of -H2A loss during ectopic recombination-mediated DSB repair 20 kb MA Ch III Ta MATa-inc Ch V -H2AX ChIP repair product Time (hr) after HO induction Note that the donor histones do not become phosphorylated (kinases may be “off” before the donor arrives)
A survey of 9 nuclear phosphoserine phosphatases reveals the PP4C phosphatase, Pph3, specifically dephosphorylates g-H2AX undamaged cells g-H2AX stain g-H2AX persists in pph3D after MMS treatment Nevan Krogan, Michael Keogh, Jeff Fillingham, Snow Shen, Andrew Emili
P P P P P P P P Wild type pph3 g-H2AX at 20 kb g-H2AX at 20 kb This suggests that g-H2AX-containing H2A/H2B dimers are displaced and subsequently dephosphorylated
Grad students Jung-Ae Kim Farokh Dotiwala Suvi Jain John Lydeard Wade Hicks Susannah Gordon-Messer Postdocs Jake Harrison Jin Li Neal Sugawara Miyuki Yamaguchi Rotation Projects in Mechanisms of DNA repair nonhomologous end-joining homologous recombination Regulation of chromosome architecture donor preference DNA damage checkpoint adaptation recovery chromatin modifications Your name here
P P P P Cohesin loading Effect of a DSB on Cohesin loading with Elcin Ünal and Doug Koshland, and Michael Lichten Smc3 Smc1 Mcd1 Scc3 will cohesin rings fall off a broken chromosome?