Download
1 / 52
650 likes | 1.49k Views
Introduction: The building blocks of the cytoskeleton Actin, actin dynamics, and actin-binding proteins Intermediate filaments. Myosins motor proteins Myosins-powered movements Actin-directed movement Cell migration. Chapters 17 and 18. Microtubules

E N D
Introduction: The building blocks of the cytoskeleton Actin, actin dynamics, and actin-binding proteins Intermediate filaments Myosins motor proteins Myosins-powered movements Actin-directed movement Cell migration Chapters 17 and 18 • Microtubules • Kinesins and dyneins; structure and function • Mitosis
The cytoskeleton of all eukaryotic cells is composed of at least two networks of filaments
Microfilaments regulate: • Cell shape • Cell movement Microtubules effect: • Localization and transport of organelles • Cell division Intermediate filaments effect: • The mechanical stability of the plasma- and the nucleus-membranes • Cell-cell interaction Accessory proteins
Cytoskeletal proteins form polymers/fibers built from small protein subunits that are held together by noncovalent bonds. • Microfilaments (F-actin): G-actin • Microtubules: aand b-tubulin • Intermediate filaments: Five subfamilies; the proteins vary in size and sequence
Small molecules can diffuse rapidly within the cytoplasm The rate of assembly of subunits into polymers, the site of assembly, and the stability of the polymer, once formed, are tightly regulated events.
In a single filament, the strength of the bond that holds subunits together is equal throughout the filament The weak side-to-side bonds are enhanced in a multi-filament complex
Actin • Highly abundant globular protein; actin comprises 1-5% of the total cellular proteins • A cell may contain half a billion actin molecules representing a cytosolic concentration of 0.5 mM • Actin is a 40 kDa protein (375 AA residues) • The protein is highly conserved in evolution; actin is encoded by multiple genes (6 in humans) that give rise to numerous isoforms. • An actin homologue, MreB, is expressed in prokaryotes
Structure of monomeric G-actin (ATP-binding cleft points towards the minus end )
The crystal structure of uncomplexed actin in the ADP stateOtterbein LR et al. Science 293:708-711, 2001 ATP hydrolysis and the release of the phosphate group results in one segment of actin switching from a strand to a helical conformation. This conformational change impacts protein-protein interactions.
Models of F-actin structure 2003 model based on fiber-diffraction data Kudryashov, Dmitry S. et al. (2005) Proc. Natl. Acad. Sci. USA 102, 13105-13110 Two parallel protofilaments wound around each other
+ + + F-actin has structural and functional polarity Experimental demonstration of polarity of an actin filament by binding of myosin S1 head domains. Myosins bind to actin with a slight tilt. When all subunits are bound by myosin, the filaments appear decorated with arrowheads that all point towards one end of the filament. This end is referred to as the “pointed end”. + _ _ Figure 18-3
The number of monomers that are added to the polymer per second is proportional to the concentration of free subunits. As the polymer grows,the concentration of free subunits drops until it reached a constant value, called the critical concentration Cc=koff/kon
The two ends of a myosin-decorated actin filament grow unequally
“Treadmilling” Actin polymerization at the leading edge of moving cells 0.12 mM 0.6 mM
How is the polymerization of actin filaments regulated? Time: 0 min Time: 15 min Cell spreading is driven by the polymerization of actin filaments
How is the polymerization of actin filaments regulated? • G-actin binding proteins • Polymerization inhibitor/Sequestering • Facilitators of actin nucleation • F-actin binding proteins • Capping proteins • Actin depolymerization factors • Severing proteins
How can we trigger and/or accelerate the formation of actin filaments? 1) Increase the availability of G-actin (intracellular concentration of G-actin is 0.5mM) 2) Increase the rate of exchange of ADP to ATP on G-actin monomers 3) Facilitate nuclei formation
Actin-binding proteins regulate the rate of assembly and disassembly, as well as the availability of G-actin for polymerization
In a platelet, the concentration of unpolymerized actin is 0.22 mM. The Cc for polymerization is 0.12 mM. How does the cell maintain such a large pool of unpolymerized actin? Thymosin b4 binds to the actin-binding region of ATP-G-actin ADP ATP (minus end) ATP Profilin Binding interrupted by PIP2 Profilin binds to proline-rich sequences found in several membrane proteins
G-actin bound to thymosin is “unavailable” for polymerization
Twisting of actin filaments induced by cofilin (Actin Depolymerizing Factor; ADF) ADF/cofilin intercalates between subdomains 1 and 2 of two longitudinally associated actin monomers causing twisting of the filament.
B A WH1 Pr o Bee 1 V C B Pr o A WH1 Scar/WAVE V C WASP B GBD A Pr o WH1 V C N-WASP GBD A WH1 B Pr o VV C PIP2 F-Actin G-Actin Cdc42 SH3 Arp 2/3 X (Grb2, Nck, IcsA Fgr, Fyn) WASP family proteins :activators of Arp2/3 complex Rac Pantaloni and Carlier Science 292:708, 2001 (Miki et al., 1998; Machesky and Insall, 1998; Rohatgi et al., 1999; Egile et al., 1999; Kim, Rosen et al., 2000; Prehoda et al;, 2000)
N-WASP mediated actin elongation and branching Pantaloni and Carlier Science 292:708, 2001
The actin severing protein, gelsolin Severed filaments serve as “nuclei” The activity of gelsolin is regulated by Ca+2 and PIP2. “strain”
Actin capping proteins block assembly and disassembly of actin filaments Limits growth Cap Z is a plus end capping protein Tropomodulin is a minus end capping protein Prevents disassembly
+Capping proteins + Gelsoin Actin polymerization is co-regulated by multiple proteins Severing protein +Cofilin
Coordinated treadmilling of distinct arrays of actin filaments in cell motility Carlier, M.-F. et al. J. Biol. Chem. 2007;282:23005-23009
Actin binding proteins regulate the rate of actin polymerization • G-actin binding proteins Thymosin b4 (inhibits polymerization) Profilin (accelerates polymerization) (PIP2) • Capping proteins Cap Z (+) (PIP2) Tropomodulin (-) • Severing proteins Gelsolin (severs and caps (+) ends)) (PIP2; Ca+2) Cofilin (ADF) (actin depolymerizing factor) (PIP2)
PIP2 affects the cytoskeleton organization
Structure/Function analysis of the interaction of PIP2 with actin-capping protein Kim et al JBC 282:5871-5879, 2007
Toxins that affect actin dynamics:(natural toxins produced by plants, sponges, and fungi) • Cytochalasin D (fungal toxin): Binds to the (+) end of actin filaments; prevents addition of subunits to existent filaments and induces depolymerization • Latrunculin (red sea sponge): Binds to G-actin and prevents its addition to filaments • Phalloidin (mushroom): Prevents depolymerization of existent filaments
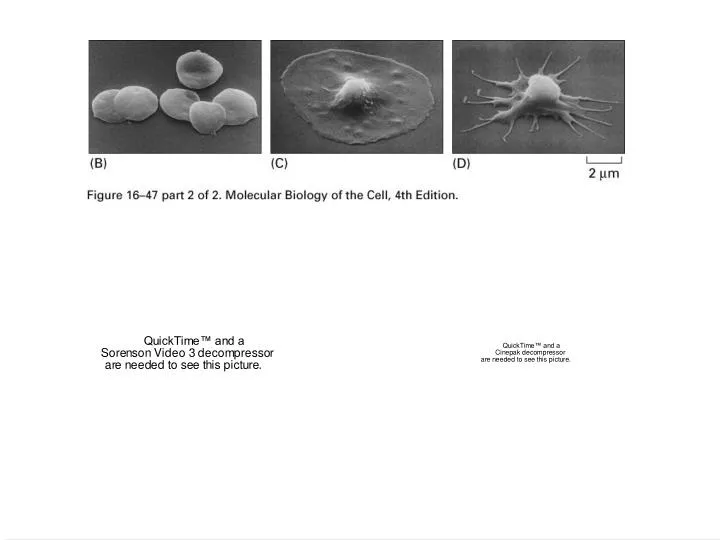
Micrograph of a detergent extracted spread platelet Micrograph of an intestinal cell showing microvilli How are the actin filaments organized spatially?
The calponin homology (CH) domain family of actin binding proteins F-actin binding domain
Chapters 17 and 18 • Introduction: The building blocks of the cytoskeleton • Actin, actin dynamics, and actin-binding proteins • Intermediate filaments
Distribution of keratins and lamins in an epithelial cell Illustration depicting the various proposed interactions of lamins with inner nuclear membrane proteins, nuclear pores, and various other nucleoplasmic factors Robert D. Goldman et al. Genes Dev. 2002; 16: 533-547
Disruption of keratin networks causes blistering Epidermolysis bullosa simplex
Various proteins cross-link intermediate filaments and connect them to other cell structures Microtubules: red intermediate filaments:blue plectin: yellow connecting-fibers: green