Download

1 / 65
660 likes | 722 Views
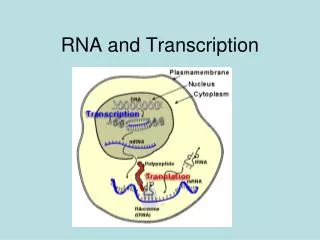
Transcription and RNA Processing. OVERVIEW OF TRANSCRIPTION The first stage in the expression of genetic information is transcription of the information in the base sequence of a ds DNA to form the base sequence of a ss RNA.

E N D
OVERVIEW OF TRANSCRIPTION • The first stage in the expression of genetic information is transcription of the information in the base sequence of a ds DNA to form the base sequence of a ss RNA. • For any particular gene, only one strand of the DNA molecule, called the template strand, is copied by RNA polymerase. • Because RNA polymerase moves in the 3' to 5' direction along the template strand of DNA, the RNA product is antiparallel and complementary to the template. • RNA polymerase recognizes start signals (promoters) and stop signals (terminators) for each of the thousands of transcription units in the genome of an organism.
Figure 1-3-1. Transcription of Several Genes on a Chromosome
TYPES OF RNA RNA molecules play a variety of roles in the cell. The major types of RNA are: • Ribosomal RNA (rRNA), which is the most abundant type of RNA in the cell. • Transfer RNA (tRNA), which is the second most abundant type of RNA. • Messenger RNA (mRNA), which carries the information specifying the amino acid sequence of a protein to the ribosome. Messenger RNA is the only type of RNA that is translated. The mRNA population in a cell is very heterogeneous in size and base sequence, as the cell has essentially a different mRNA molecule for each of the thousands of different proteins made by that cell.
Heterogeneous nuclear RNA (hnRNA or pre-mRNA), which is found only in the nucleus of eukaryotic cells. It represents precursors of mRNA, formed during its posttranscriptional processing. • Small nuclear RNA (snRNA), which only functions in the nucleus of eukaryotes. One of its major functions is to participate in splicing (removal of introns) mRNA. • Micro-RNA**: short, non-coding, ~ 22 nt long, at least some of which control the expression or repression of other genes during development. • Ribozymes**, which are RNA molecules with enzymatic activity. They are found in both prokaryotes and eukaryotes.
TRANSCRIPTION: IMPORTANT CONCEPTS AND TERMINOLOGY • RNA polymerase locates genes in DNA by searching for promoter regions. • The promoter is the binding site for transcription factors and RNA polymerase. • Binding establishes where transcription begins, which strand of DNA is used as the template, and in which direction transcription proceeds.
RNA polymerase moves along the template strand in the 3' to 5' direction as it synthesizes the RNA product in the 5' to 3' direction using NTPs (ATP, GTP, CTP, UTP) as substrates. RNA polymerase does not proofread its work. The RNA product is complementary and antiparallel to the template strand. • The coding (non-template) strand is not used during transcription. It is identical in sequence to the RNA molecule, except that RNA contains uracil instead of the thymine found in DNA. • By convention, the base sequence of a gene is given from the coding strand (5' 3').
In the vicinity of a gene, a numbering system is used to identify the location of important bases. The first base transcribed as RNA is defined as the + 1 base of that gene region. To the left (5', or upstream) of this starting point for transcription, bases are -1, -2, -3, etc. to the right (3', or downstream) of this point, bases are +2, +3, etc. • Transcription ends when RNA polymerase reaches a termination signal.
Figure 1-3-3. Flow of Genetic Information From DNA to Protein
Sample Questions 1. During RNA synthesis, the DNA template sequence TAGC would be transcribed to produce which of the following sequences? A. ATCG B. GCTA C. CGTA D. AUCG E. GCUA
2. Transcription of the following sequence of the tryptophan operon occurs in the direction indicated by the arrow. What would be the base sequence of the mRNA produced? 3'... CGCCGCTGCGCG... 5' 5'... GCGGCGACGCGC. . .3' A. 5'...GCGGCGACGCGC...3' B. 5'...GCGCGUCGCCGC...3' C. 5'...GCGCGTGCGGCG...3' D. 5'...GCGGCGUCGCGC...3' E. 5'... CGCGCTCGCCGC...3'
RNA POLYMERASES • There is a single prokaryotic RNA polymerase that synthesizes all types of RNA in the cell. • The core polymerase responsible for making the RNA molecule has the subunit structure (α2ββ`). • A protein factor called sigma (σ) is required for the initiation of transcription at a promoter. Sigma factor is released immediately after-initiation of transcription. • Termination of transcription sometimes requires a protein called rho (ρ) factor. • This enzyme is inhibited by rifampin. • Actinomycin D binds to the DNA preventing transcription.
Promoter “Strength” Affects Amount of RNA made, Thus it Affects level of Expression for that Gene. Not all promoters have same “strength” (activity)! • Promoters differ in DNA sequences and “strength” • RNA polymerase binds differently to different sequences • “Strong promoters” initiate transcription more often than “weak promoters” • rRNA has strong promoter: ~1 initiation per second • lacZ has a weak promoter: ~1 initiation per minute
There are three eukaryotic RNA polymerases, distinguished by the particular types of RNA they produce: • RNA polymerase I is located in the nucleolus and synthesizes 28S, 18S, and 5.8S rRNAs. • RNA polymerase II is located in the nucleoplasm and synthesizes hnRNA/mRNA, miRNA and some snRNA. • RNA polymerase III is located in the nucleoplasm and synthesizes tRNA, some snRNA, some nc-RNA, and 5S rRNA.
Transcription factors (such as TFIID for RNA polymerase II) help to initiate transcription. • The requirements for termination of transcription in eukaryotes are not well understood. • All transcription can be inhibited by actinomycin D. • In addition, RNA polymerase II is inhibited by (α-amanitin) a toxin from certain mushrooms.
Alpha-Amanitin Strongly Inhibits RNA Pol II • Alpha-amanitin is strong inhibitor (toxin) from “death cap” • Inactivates RNA pol II and can kill a person • RNA pol I and III are less affected by toxin
Comparison of Eukaryotic and Prokaryotic RNA Polymerases miRNA
The following events occur during the expression of a prokaryotic gene: • With the help of sigma factor, RNA polymerase recognizes and binds to the promoter, region. • The bacterial promoter contains two "consensus" sequences, called the Pribnow box (or TATA box) and the -35 sequence. The promoter identifies the start site for transcription and orients the enzyme on the template strand. 2. Transcription begins at the + 1 base pair. Sigma factor is released as soon as transcription is initiated. 3. The core polymerase continues moving along the template strand in the 3' to 5' direction, synthesizing the mRNA in the 5' to 3' direction.
4. RNA polymerase eventually reaches a transcription termination signal, at which point it will stop transcription and release the completed mRNA molecule. There are two kinds of transcription terminators commonly found in prokaryotic genes: • Rho-independent termination occurs when the newly formed RNA folds back on itself to form a GC-rich hairpin loop closely followed by 6-8 U residues. These two structural features of the newly synthesized RNA promote dissociation of the RNA from the DNA template. • Rho-dependent termination requires participation of rho factor. This protein binds to the newly formed RNA and moves toward the RNA polymerase that has paused at a termination site. Rho then displaces RNA polymerase from the 3' end of the RNA.
5. Transcription and translation can occur simultaneously in bacteria. Because there is no processing of prokaryotic mRNA (generally no introns), ribosomes can begin translating the message even before transcription is complete. - Ribosomes bind to a sequence called the Shine-Dalgarno sequence in the 5' untranslated region (UTR) of the message. - Protein synthesis begins at an AUG codon at the beginning of the coding region and continues until the ribosome reaches a stop codon at the end of the coding region.
6. The ribosome translates the message in the 5' to 3' direction, synthesizing the protein from amino terminus to carboxyl terminus.
The mRNA produced by the gene shown above is a monocistronic message. That is, it is transcribed from a single gene and codes for only a single protein. • The word cistron is another name for a gene. • Some bacterial operons produce polycistronic messages. In these cases, related genes grouped together in the DNA are transcribed as one unit. • The mRNA in this case contains information from several genes and codes for several different proteins
Prokaryotic Polycistronic Message Codes for Several Different Proteins
PRODUCTION OF EUKARVOTIC MESSENGER RNA • In eukaryotes, most genes are composed of coding segments (exons) interrupted by noncoding segments (introns). • Both exons and introns are transcribed in the nucleus. • Introns are removed during processing of the RNA molecule in the nucleus. • In eukaryotes, mRNA is usually monocistronic. The mature mRNA is translated in the cytoplasm.
Human Genome Organization is complex • Gene density varies enormously from chromosome to chromosome and within different regions of the same chromosome. • In chromosomal regions with high gene density, overlapping genes may be found; they are typically transcribed from opposing DNA strands. • Sometimes the overlaps are partial, but in other cases small protein-coding genes are located within the introns of larger genes. • Recent analyses have also shown that ncRNA genes can frequently overlap protein-coding genes.
Transcription of a typical eukaryotic gene occurs as follows: • With the help of proteins called transcription factors, RNA polymerase II recognizes and binds to the promoter elements. 2. RNA polymerase II separates the strands of the DNA over a short region to initiate transcription and read the DNA sequence. The template strand is read in the 3' to 5' direction as the RNA product (the primary transcript) is synthesized in the 5' to 3' direction. Both exonsand introns are transcribed.
Class II Promoters (Most similar to Bacterial promoters) Common type of promoter (most genes use this) • Many variations, but “consensus” has a “Core” + “Upstream” • Core promoter elements (motifs): contribute to basal transcription 1. TFIIB recognition element (BRE) 2. “TATA box” (5’-TATA-3’) 3. Initiator box (Inr) with an “A” at +1, most common 4. Motif ten element (MTE) 5. Downstream promoter element (DPE, less common) • A particular core promoter may contain some, all, or none of these motifs • Core promoter is recognized by general TFs that associate with RNA pol to form a pre-initiation complex at great majority of promoters
Class II Promoters Cont. • Proximal promoter - the proximal sequence upstream of the gene that tends to contain primary regulatory elements • Approximately 250 base pairs upstream of the start site • Generaltranscription factor binding sites • Distal promoter - the distal sequence upstream of the gene that may contain additional regulatory elements • Enhancers • Silencers • Locus control region • Insulators • Specific transcription factor binding sites
Examples of Proximal Promoter Elements: • GC box (GC rich) • CAAT box (5’-CCAAT-3) • e.g., GC boxes bind the TF Sp1, while CCAAT boxes bind CTF
3. RNA polymerase II ends transcription when it reaches a termination signal. One Model: • The eukaryotic RNA polymerase II usually continues to transcribe well past the end of the gene. • After the end of the gene has been reached, RNA polymerase II passes through one or more AATAAA sequences, which lie beyond the 3' end of the coding • The pre-mRNA, carrying this signal as AAUAAA, is then cleaved by a special endonuclease that recognizes the signal and cuts at a site 11 to 30 residues to its 3' side. • A tail of polyriboadenylic acid, poly(A), as much as 200 bases long, is added by a special non-template-directed polymerase.
Transcription Enhancers and Silencers • Both are Binding Sites for Transcription Factors (TF’s) • Enhancers: Increase the amount of Transcription from a nearby promoter (core + proximal promoter elements) • Silencers: Decrease amount of Transcription from nearby promoters • Initially Defined as being “Position and orientation independent” • Found upstream, within, or downstream of genes • Function in either orientation (not always true)
What Are Enhancers? Enhancer = “non-promoter DNA elements that stimulate transcription” • First found in eukaryotes and then found in bacteria • Binding sites for transcription factors (= activators or enhancer binding proteins). These proteins interact with general transcription factors to promote formation of pre-initiation complex. • NOT the binding sites for RNA polymerase • Can function over long distances (100 - 1000 bp) away from +1 sites (upstream or downstream) • Orientation not important (can be flipped and still work) • classical way to distinguish enhancer elements • Many are also tissue-specific (rely on tissue-specific DNA-binding proteins for activity). • Sometimes a DNA element can act as an enhancer or a silencer depending on what is bound to it.
TATA Box By Itself is a Weak Promoter • Additional promoter sequences (such as GC, GAL4 etc…) are needed for a strong promoter OR for regulated promoters • These additional regulatory / enhancer sequences often act as binding sites for Gene-specific Transcription Factors Holo-TFIID can respond to a wide variety of activators, represented here by Sp1, Gal4-NTF-1, and a generic activator (green) at top.
Processing of Eukaryotic Messenger RNA The primary transcript must undergo extensive posttranscriptional processing inside the nucleus to form the mature mRNA molecule. These processing steps include: 1. A 7-methylguanosine cap is added to the 5' end while the RNA molecule is still being synthesized. The cap structure serves as a ribosome-binding site and also helps to protect the mRNA chain from degradation.
2. A poly-A tail is attached to the 3' end. In this process, an endonuclease cuts the molecule on the 3' side of the sequence AAUAAA (poly-A addition signal), then poly-A polymerase adds the poly-A tail (about 200 As) to the new 3' end. The poly-A tail protects the message against rapid degradation and aids in its transport to the cytoplasm. A few mRNAs (for example, histone mRNAs) have no poly-A tails.
3. Introns are removed from hnRNA by splicing, accomplished by spliceosomes (also known as an snRNP, or snurp) , which are complexes of snRNA and protein. The hnRNA molecule is cut at splice sites at the 5' (donor) and 3' (acceptor) ends of the intron. - The intron is excised in the form of a lariat structure and degraded. Neighboring exons are joined together to assemble the coding region of the mature mRNA.
Nuclear Splicing Signals (RNA sequences) that Define Introns • Consensus sequences for exon/intron boundaries known • “GU-AG rule” = most nuclear introns can be defined by these sequences • Branch site is within intron and has critical “A” nt
Spliceosome catalyzes two Trans-esterification Reactions 1) 2` OH on the “A” in the branch site “attacks” the “Phosphate” on “G” at the 5` splice site - This creates an unusual 2’ -5’ branched nucleic acid (“lariat”) 2) 3’ OH at the exposed 5’ splice site “attacks” the “Phosphate” on the “G” at the 3’ splice site Result: - covalently linked exons - formation of lariat intron, degraded
4. All of the intermediates in this processing pathway are collectively known as hnRNA. 5. The mature mRNA molecule is transported to the cytoplasm, where it is translated to form a protein. Note Mutations in splice sites can lead to abnormal proteins. For example, mutations that interfere with proper splicing of β-globin mRNA are responsible for some cases of β-thalassemia.