Download

1 / 21
210 likes | 227 Views
Discover the processes behind adaptive Macroevolution through positive selection, allele replacements, inbreeding depression, and more in natural populations.

E N D
Topic 15. Lecture 21. Microevolutionary mechanisms of Macroevolution. We already considered genetic variation of natural populations and factors that affect this variation (separately and together). Finally, we are prepared to address the key question: what happens in natural populations? We will concentrate on changes driven by positive selection, as they are behind adaptive Macroevolution. However, most of genetic variation present in natural populations is irrelevant for adaptive Macroevolution, and we need to briefly consider such variation, too. Specifically, we will treat the following subjects: i) How strong could natural selection be in natural populations? ii) Inbreeding depression and outbreeding depressions - two pervasive phenomena. iii) How common are selectively neutral variation and allele replacements? iv) How common are unconditionally deleterious alleles, which will never be fixed? v) Microevolution of extinction. vi) How common are positive selection-driven allele replacements? vii) How Microevolution constraints the rate and the course of Macroevolution?
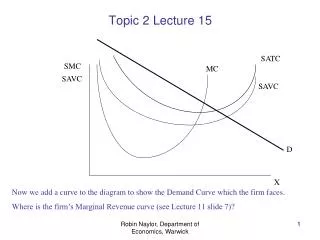
i) How strong could natural selection be in natural populations? 1) Is natural selection consistent with survival of the population? All kinds of natural selection are involved with some genetic load. Thus, if the maximal contribution of an individual to the next generation is one offspring (two for sexual diploids), there could be no selection - any selection will drive the population to extinction. However, in every population, there is an opportunity for selection, because maximal fecundity is always higher than 2 - apparently, it is never below 10 (great apes, some birds of prey). Also, contributions of males usually vary greatly, creating more opportunity for selection. Apparently, any natural population can take a genetic load of at least 50%. 2) Is natural selection operating within natural populations? It is hard to measure genetic load experimentally, but it can be done for variance of relative fitness Var[wi/W]. This quantity can be referred to as "opportunity for selection": if Var[wi/W] = 0, there could be no selection. However, data consistently show that fitness and its components are variable within natural populations. Opportunity for selection on some fitness-related traits in Drosophila melanogaster Fecundity 0.15 Longevity 0.07 Developmental time 0.002 All natural populations, except abnormally small ones, possess a lot of non-neutral genetic variation. Of course, not all of this variation may be used for adaptive evolution. Traits that are closely related to fitness (viability, fecundity, mating success, longevity) have highest evolvabilities (Genetics 130, 195-204, 1992).
Sexual selection creates extra opportunity for selection among males, because often a small fraction of males obtain most of matings. Number of mating partners of an individual in a natural population of green swordtails (Molecular Ecology 17, 4522-4534, 2008).
ii) Inbreeding depression and outbreeding depressions - two pervasive phenomena. A) Inbreeding depression. Offspring from matings between closely related parents consistently have reduced fitnesses. This inbreeding depression increases with the coefficient of inbreeding F, the fraction of allele, at the same locus, that are "identical-by-descent", in the sense so they can be traced to the common ancestor. Inbreeding depression has been detected in almost all experiments that studied the impact of inbreeding on fitness. Traits that are closely related to fitness (viability, fecundity, mating success, longevity) display the largest values of inbreeding depression. Very roughly, the increase of F by 1% cause the decline of a fitness-related trait that is also ~1%. Thus, complete homozygosity leads to fitness that is close to 0. This is not surprising, as long as an average individual carries ~1-2 heterozygous recessive lethals. In fact, inbreeding depression is caused not only by recessive lethals, but also by deleterious alleles with individually mild effects.
Inbreeding depression of fertility in a tree-hole-breeding mosquito Aedes geniculatus(Proc. Royal Society B, 267, 1939 - 1945, 2000).
Inbreeding depression may depend on the environment under which fitness is assayed. Not surprisingly, it is often higher under tough, competitive environments. The reproductive success of inbred (solid bars) and outbred (open bars) mice. (A) Males in enclosures. (B) Males in laboratory (PNAS 97, 3324-3329, 2000). Male mice are really mean!
Naturally, inbreeding increases the fraction of homozygous loci in offspring, and this probably is the cause of inbreeding depression. Still, there are two (not mutually exclusive) feasible explanations of inbreeding depression - dominance and overdominance. Overdominance (red): Aa has the highest fitness, and, thus, homozygosity is always bad. Dominance of beneficial alleles (blue): one of homozygotes (aa) has reduced fitness, and, thus, homozygosity is bad on average. Overdominance is rare, so that dominance (incompatibility epistasis between deleterious alleles) is probably the leading cause of inbreeding depression.
B. Outbreeding depression. Matings between close relatives lead to low fitness of offspring, but matings between parents from different populations (from "the same species") may lead to the same effect - and this is called outbreeding depression. Mass of individual seeds versus pollination distance in the marine angiosperm Zostera marina (Marine Biology 152, 793-801, 2007). Thus, often there is an intermediate optimal genetic distance between parents. As inbreeding depression, outbreeding depression is due to incompatibility epistasis, but between alleles of different loci. We will return to this subject when we consider speciation.
iii) How common are selectively neutral variation and allele replacements? In short, very common. The highest values of H are present at sites that are under no or only very weak selection. In compact genomes of prokaryotes and of some eukaryotes, such as Drosophila melanogaster (Nature 437, 1149-1152, 2005), such sites are rare, but they represent at least 80% of mammalian genomes. An average human carries ~5,000,000 (effectively) neutral variants (with s < 1/Ne < 10-5), most of them rare (H = 10-3, diploid genome size = 6x109). Because neutral alleles at a locus become fixed at a rate that equals to mutation rate, the total number of selectively neutral allele replacements per generation was ~50 in the human lineage. That is how humans and chimpanzees accumulated ~50,000,000 genetic differences (mostly, single-nucleotide replacements) in the course of only ~1,000,000 generations of independent evolution. In nonmammalian species, these numbers may be lower, but still impressive. Thus bulk of genetic differences between species is due to (effectively) neutral allele replacements. Indeed, it is hard to imagine positive selection that drives 50 adaptive allele replacements every generation (although theoretically this might be feasible).
iv) How common are unconditionally deleterious alleles, which will never be fixed? Also quite common, because a lot of such alleles are only mildly deleterious - this is enough to prevent them from being fixed, but not enough to keep them very rare. An average Drosophila, fish (and probably human) carries 1-2 heterozygous recessive lethals, which reduce fitness by ~1%. An average normal human carries at least ~30 deleterious alleles that completely inactivate the affected proteins (nonsense substitutions, frameshift insertions and deletions, etc.), but still are not recessive lethals, because these proteins are not essential. Coefficients of selection against such alleles are ~10-2-3. An average "normal" human carries at least ~1000 nonsynonymous mildly deleterious variants (out of ~10,000 nonsynonymous variants totally) with coefficients of selection against them ~10-3-4(Hum. Mol. Genet. 10, 591-597 2001). Perhaps, a number of mildly deleterious non-coding alleles is even higher. However, almost all drastic deleterious alleles affect protein sequences. Data for other eukaryotes are probably similar. Humans are just better studied. So, only ~10% of human non-synonymous variants are substantially deleterious, having selection coefficients above 10-5, at the very least. However, we know that ~90% of human non-synonymous mutations are substantially deleterious. Obviously, deleterious allele are underrepresented in within-population variation, due to negative selection.
Probabilities that an amino acid replacement is associated with the coefficient of selection s > 10–2, 10–2–10–4, 10–4–10–5 or < 10–5. Hum. Mol. Genet. 14, 3191-3201, 2005). Grey: probabilities among fresh mutations. Red: probabilities among segregating alleles. Neutral and mildly deleterious variants are overrepresented. Thus, the bulk of genetic variation within every population is either irrelevant for adaptive evolution (effectively neutral variation) or has not chance to become fixed within an evolving lineage (unconditionally deleterious variation). Nature is wasteful, as usual.
v) Microevolution of extinction. In fact, there is one aspect of Microevolution that is directly affected by deleterious mutations and negative selection - extinction. Extinction is a complex phenomenon - unless it is caused by a complete habitat destruction or by direct killing of every individual ("hard extinction"). A soft extinction involves an ecological component: 1) critical low density (Allee effect) - it could be that if too few individuals are left, they cannot reproduce 2) demographical stochasticity - in a small population there is a substantial chance that the number of individuals will decline further "by chance" and a genetical component: 1) inbreeding depression due to homozygosity of pre-existing deleterious alleles 2) fixation of new deleterious mutations due to inefficient selection under low Ne 3) loss of genetic variation, which prevents response to positive selection Genetic deterioration of ecologically declining populations plays an important role in soft extinctions. Low population size leads to genetic deterioration, and vice versa, leading to a self-reinforcing process known as mutational meltdown. Two kinds of data demonstrate the importance of genetic deterioration in extinction.
1) Very small populations on the verge of extinction often contain genotypes of low quality, presumably due to fixation of mildly deleterious alleles. This can be seen by the results of crosses of such genotypes with members of a related, "healthy" population, if such a population is available. South Florida houses a small, isolated, inbred and distinct subspecies of panthers (Puma concolor coryi). Once part of a continuous, widespread population, panthers became isolated in south Florida more than a century ago. Numbers declined and the occurrence of genetic defects increased. Hoping to reverse the genetic damage, managers introduced eight female panthers from Texas into south Florida in the mid-1990s. Hybrid kittens have about a three times higher chance of becoming adults as do purebred ones. Hybrid adult females survive better than purebred females; there is no obvious difference between the males. Hybrids are expanding the known range of habitats panthers occupy and use (Animal Conservation9, 115-122, 2006). The Florida panther Puma concolor coryi .
2) If natural selection is artificially abolished, mean population fitness declines rapidly, by 1-2% per generation, due to unopposed accumulation of new deleterious mutations. This can be seen from data on "Middle Class Neighborhood" (MCN) populations and on reintroductions of individuals from captive populations into the wild. An MCN population: if every pair contributes exactly two offspring to the next generation, natural selection is impossible (although genetic drift still occurs slowly). In an MCN population, fitness declines by 1-2% per generation.
Decline of fitness, measured under tough conditions, in populations of different sizes maintained in captivity under relaxed natural selection (Proc. Natl. Acad. Sci. USA 94, 13034-13039, 1997; Conservation Genetics 3, 277–288, 2002; Biological Conservation 126, 131-140, 2005; Ann. Rev. Ecol. Evol. Syst. 37, 433-458 2006).
vi) How common are positive selection-driven allele replacements? Finally, we can consider the only aspect of Microevolution that is directly responsible for adaptive Macroevolution - allele replacements driven by positive selection. There are two main questions: 1) What is the per generation number of number of selection-driven allele replacements accepted by an evolving population? 2) What are coefficients of selection that drives these replacements? Due to difficulties in detecting positive selection, we still do not have definite answers to these fundamental questions. However, there are some estimates, for a small number of carefully studied species, first of all for several species of Drosophila and for humans.
Positive selection in Drosophila melanogaster lineage. D. melanogaster is originally an African species that dispersed into other parts of the Old World ~15,000 years ago. Since the divergence of African and non-African D. melanogaster, there were ~200 selection-driven allele replacements in Africa and ~300 such replacements outside Africa. Assuming 20 generations per year, this implies 1 adaptive allele replacement per 1000 generations. Most of these replacements were driven by selection with coefficients 0.05%-0.5% (PLoS Genet 2, e166, 2005). I tend to believe this estimate, although I am not 100% sure. Perhaps, adaptive allele replacements are more common, but hardly more than 1 per 100 generations (Trends in Ecology & Evolution 21, 569-575, 2006).
Positive selection in the human lineage. As we know, it must be easier to detect recent or even ongoing episodes of positive selection, through the pattern of genetic variation. Extensive data on genetic variation within humans are available, but so far they did not translate into any credible quantitative estimates, although several cases, such as lactose tolerance, are well established. What we have now is a mess. This table demonstrates discrepancies in estimates of positive selection in humans. For each pair of studies, is presents the number of genes that were identified as being under positive selection in both of them (Nature Reviews Genetics 8, 857-868, 2007). It seems very likely, however, that the average rate of selection-driven allele replacements in human evolution was less than 1 replacement per 100 generations, but we do not know how much less, and there are no credible estimates of the strengths of selection involved.
Putative conclusions about positive selection: 1) A vast majority of non-neutral genetic variation within every population is totally irrelevant to its evolution - it is due to segregation of rare, derived, deleterious alleles that will never be fixed. 2) A vast majority of allele replacements is irrelevant to adaptive evolution - these replacements involve selectively neutral (or almost neutral) pairs of alleles. 3) Allele replacements driven by substantial positive selection are not very common (1 per 100 or 1000 generations) and, thus, probably overlap only very little. Still, we have no idea, how many adaptive allele replacements are responsible for the transition from humans to apes. How many chimpanzee alleles one need to replace to turn it into a passable human? 100? 1000? 10000? 10000?
vii) How Microevolution constraints the rate and the course of Macroevolution? Darwinian mechanism of Microevolution is very inefficient, in comparison with imaginary Lamarckian mechanism. Thus, it makes sense to ask the above question. There is no comprehensive answer, but we probably can say the following: 1) Microevolutionary mechanism prevent long-term survival of lineages represented by too few individuals at a time. At least 1,000 or even 10,000 individuals are needed. 2) The course of Macroevolution is severely constrained by availability of mutations - an insertion of a particular sequence of 10 amino acids into the protein may never occur. 3) The rate of Macroevolution is limited by availability of even relatively common mutations: a population of 100,000 individuals will need to wait for 100,000 generations before a nucleotide substitution will occur with probability 10-8 and survive the initial random drift with probability 10-2 (assuming 1% selective advantage). 4) Fixations of individual mutations make Macroevolution unpredictable. This is a huge magnification of quantum effects. 5) It does not seem that inherent inefficiency of natural selection imposes a strong constraint on Macroevolution.
Quiz: What features of a population and of its environment can be expected to cause a high rate per generation number of adaptive allele replacements in it? In particular, who should accumulate beneficial mutations faster, fruit flies or humans? Hint: think of possible effects of mutation rate, population structure, drift, and changes of external conditions.