Download
1 / 1
10 likes | 277 Views
Role of isoforms of chalcone synthase in controlling flux into flavonoid pathways Alexandra Lanot* , Steven P Colliver, Mark P Robbins and Philip Morris. 1. 3. 2. Lotus japonicus , a convenient model to study flux between flavonoid pathways.

E N D
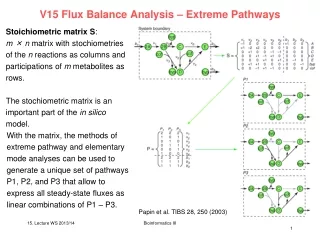
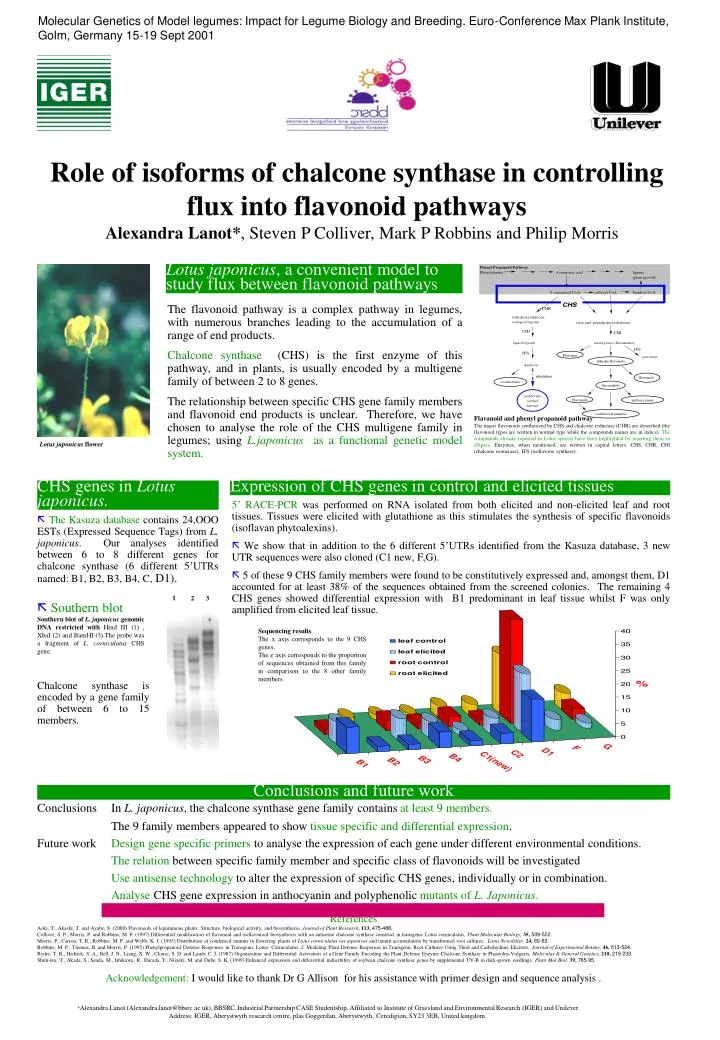
Role of isoforms of chalcone synthase in controlling flux into flavonoid pathways • Alexandra Lanot*, Steven P Colliver, Mark P Robbins and Philip Morris 1 3 2 Lotus japonicus, a convenient model to study flux between flavonoid pathways The flavonoid pathway is a complex pathway in legumes, with numerous branches leading to the accumulation of a range of end products. Chalcone synthase (CHS) is the first enzyme of this pathway, and in plants, is usually encoded by a multigene family of between 2 to 8 genes. The relationship between specific CHS gene family members and flavonoid end products is unclear. Therefore, we have chosen to analyse the role of the CHS multigene family in legumes; using L.japonicus as a functional genetic model system. Flavanoid and phenyl propanoid pathway The major flavonoids synthesised by CHS and chalcone reductase (CHR) are described (the flavonoid types are written in normal type while the compounds names are in italics). The compounds already reported in Lotus species have been highlighted by inserting them in ellipses. Enzymes, when mentioned, are written in capital letters: CHS, CHR, CHI (chalcone isomerase), IFS (isoflavone synthase). Lotus japonicus flower CHS genes in Lotus japonicus. Expression of CHS genes in control and elicited tissues 5’ RACE-PCR was performed on RNA isolated from both elicited and non-elicited leaf and root tissues. Tissues were elicited with glutathione as this stimulates the synthesis of specific flavonoids (isoflavan phytoalexins). We show that in addition to the 6 different 5’UTRs identified from the Kasuza database, 3 new UTR sequences were also cloned (C1 new, F,G). 5 of these 9 CHS family members were found to be constitutively expressed and, amongst them, D1 accounted for at least 38% of the sequences obtained from the screened colonies. The remaining 4 CHS genes showed differential expression with B1 predominant in leaf tissue whilst F was only amplified from elicited leaf tissue. The Kasuza database contains 24,OOO ESTs (Expressed Sequence Tags) from L. japonicus. Our analyses identified between6 to 8 different genes for chalcone synthase (6 different 5’UTRs named: B1, B2, B3, B4, C, D1). Southern blot Southern blot of L. japonicus genomic DNA restricted with Hind III (1), XbaI (2) and BamHI (3).The probe was a fragment of L. corniculatus CHS gene. Sequencing results The x axis corresponds to the 9 CHS genes. The z axis corresponds to the proportion of sequences obtained from this family in comparison to the 8 other family members. Chalcone synthase is encoded by a gene family of between 6 to 15 members. Conclusions and future work Conclusions In L. japonicus, the chalcone synthase gene family contains at least 9 members. The 9 family members appeared to show tissue specific and differential expression. Future work Design gene specific primers to analyse the expression of each gene under different environmental conditions. The relation between specific family member and specific class of flavonoids will be investigated Use antisense technology to alter the expression of specific CHS genes, individually or in combination. Analyse CHS gene expression in anthocyanin and polyphenolic mutants of L. Japonicus. References Aoki, T., Akashi, T. and Ayabe, S. (2000) Flavonoids of leguminous plants: Structure, biological activity, and biosynthesis, Journal of Plant Research,113, 475-488. Colliver, S. P., Morris, P. and Robbins, M. P. (1997) Differential modification of flavonoid and isoflavonoid biosynthesis with an antisense chalcone synthase construct in transgenic Lotus corniculatus, Plant Molecular Biology,35, 509-522. Morris, P., Carron, T. R., Robbins, M. P. and Webb, K. J. (1993) Distribution of condensed tannins in flowering plants of Lotus corniculatus var japonicus and tannin accumulation by transformed root cultures., Lotus Newsletter,24, 60-63. Robbins, M. P., Thomas, B. and Morris, P. (1995) Phenylpropanoid Defense Responses in Transgenic Lotus- Corniculatus .2. Modeling Plant Defense Responses in Transgenic Root Cultures Using Thiol and Carbohydrate Elicitors, Journal of Experimental Botany,46, 513-524. Ryder, T. B., Hedrick, S. A., Bell, J. N., Liang, X. W., Clouse, S. D. and Lamb, C. J. (1987) Organization and Differential Activation of a Gene Family Encoding the Plant Defense Enzyme Chalcone Synthase in Phaseolus-Vulgaris, Molecular & General Genetics,210, 219-233. Shim-izu, T., Akada, S., Senda, M., Ishikawa, R., Harada, T., Niizeki, M. and Dube, S. K. (1999) Enhanced expression and differential inducibility of soybean chalcone synthase genes by supplemental UV-B in dark-grown seedlings, Plant Mol Biol,39, 785-95. Acknowledgement: I would like to thank Dr G Allison for his assistance with primer design and sequence analysis . *Alexandra Lanot (Alexandra.lanot@bbsrc.ac.uk), BBSRC Industrial Partnership CASE Studentship, Affiliated to Institute of Grassland and Environmental Research (IGER) and Unilever Address: IGER, Aberystwyth research centre, plas Goggerdan, Aberystwyth, Ceredigion, SY23 3EB, United kingdom. Molecular Genetics of Model legumes: Impact for Legume Biology and Breeding. Euro-Conference Max Plank Institute, Golm, Germany 15-19 Sept 2001