Download

1 / 58
670 likes | 1.31k Views
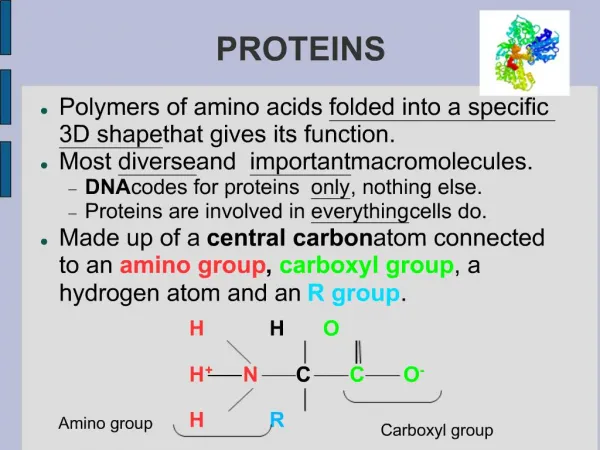
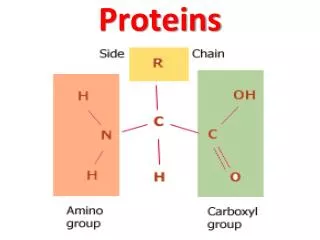
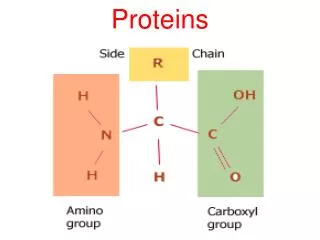
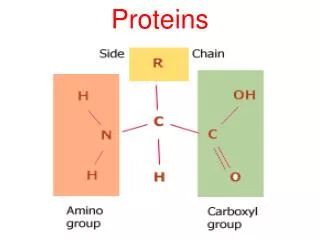
PROTEINS. i. FIBROUS PROTEINS Collagen Elastin Keratin ii. GLOBULAR PROTEINS Myoglobin Hemoglobin. COLLAGEN. STRUCTURE Triple helix Has 3.3 residues per turn and a rise per residue nearly twice that of an -helix Every 3 rd amino acid residue is a glycine residue

E N D
PROTEINS i. FIBROUS PROTEINS Collagen Elastin Keratin ii. GLOBULAR PROTEINS Myoglobin Hemoglobin
COLLAGEN STRUCTURE • Triple helix • Has 3.3 residues per turn and a rise per residue nearly twice that of an -helix • Every 3rd amino acid residue is a glycine residue • Gly-Pro-X or Gly-Hyp-X
Hydroxyproline (Hyp) • Result from hydroxylation of proline residue • Stabilizes the triple helical structure of collagen Ascorbate
m-RNA Signal protein Biosynthesis of Collagen 1. Synthesis of a chains of pre-procollagen on ribosomes. A signal protein directs them to the RER .
OH OH OH OH OH OH 2. Cleavage of signal protein forms procollagen 3. Hydroxylation of lysine and proline Lysine Hydroxylysine Peptidyl lysine hydroxylase Proline Hydroxyproline Peptidyl proline hydroxylase Ascorbic acid is necessary to activate the hydroxylases
OH OH OH OH OH OH OH 4. Glycosylation • Addition of galactose and glucose to some hydroxylysine residues. • The enzymes galactosyl transferase and glycosyl transferase are required for this process.
S S S S S S S S S S S S Registration peptides 5. Assembly of three - chains to form procollagen This involves the formation of disulfide bonds between parts of the polypeptide chains known asregistration peptides, which occur at both ends of the pre-procollagen. • The assembly process aligns the three a - chains relative to one another. • The three alpha chains are wound around one another in the form of a triple helix. • The assembly process occurs in the Golgi apparatus.
6. Secretion of procollagen molecules by exocytosis into the extra cellular space
Procollagen peptidase Procollagen peptidase 7. Cleavage of registration peptides • Occurs in the extra cellular space, and is catalysed by procollagen peptidases. • The resulting molecule is called tropocollagen.
8. Self-assembly or polymerization of tropocollagen molecules form collagen fibrils • Cross-linkage between adjacent tropocollagen molecules stabilizes the fibrils. • It involves the removal of an amino group (NH2), which has a net oxidative effect and the formation of covalent cross-links. • It is catalyzed by lysine oxidase (or catalase).
Inheritable defects in collagen molecule Characterized by stretchy skin and loose joints Due to defect in genes that encode -collagen-1, procollagen N-peptidase, or lysyl hydroxylase Ehlers-Danlos Syndrome
Osteogenesis imperfecta Brittle bone syndrome Bones easily bend and fracture COLLAGEN DISEASES
COLLAGEN DISEASES • Menkes is a genetic disorder caused by a mutation usually primarily in the Y Chromosome (boys) it affects the copper levels and metabolism in the body causing seizures, brain damage, weakened bones and muscles, organ shutdown and failure to thrive. • There is no cure and unless caught within days of birth medication and treatment is only symptomatic. • Children with the disease will die before their first decade of life, most dying within infancy or toddlerhood. Menke’s syndrome • Characterized by kinky hair and growth retardation • Due to dietary deficiency of copper required by lysyl oxidase, which catalyzes a key step in the formation of the covalent cross-links that strengthen collagen fibers. • Less than 1 in 1,250,000 children will be born with Menkes Disease, otherwise known as Kinky Hair Syndrome.
Scurvy • Best known defect in collagen biosynthesis • Deficiency of Vit.C (required by prolyl and lysyl hydroxylases) • Bleeding gums, swelling joints, poor wound healing, and ultimately death.
Elastin • Connective tissue protein with rubber-like properties • Found in lungs, walls of large blood vessels, and elastic ligaments • Can be stretched to several time their normal length, but recoil to their original shape when relaxed
STRUCTURE OF ELASTIN • Composed primarily of small non polar amino acid residues (e.g. G, A, V) • Also rich in proline and lysine, but contains little hydroxyproline and hydroxylysine • Interchain cross-links form desmosine residues
DESMOSINE CROSS-LINK • An extensively interconnected, rubbery network that can stretch and bend in any direction when stressed, giving connective tissue elasticity
Role of -1 antitrypsin in elastin degradation • Inhibit neutrophil elastase (protease that degrades elastin of alveolar walls) Deficiency of -1 antitrypsin leads to destruction of the alveolar walls of the lungs resulting to EMPHYSEMA Treatment : Administration of -1 AT
Role of -1 antitrypsin in elastin degradation • As we breath, we not only take in oxygen, but we also take in our environment. • Pollen, sawdust, car exhaust, dander, paint fumes and over-spray, radon gases, hair sprays, perfumes, household cleansing fumes, tobacco smoke and many other pollutants
Role of -1 antitrypsin in elastin degradation • Elastase - the lungs natural defense against the irritants • Elastase (shown as white dots) attach themselves to the foreign material in the sac and consumes it as well as bacteria in the lungs
Role of -1 antitrypsin in elastin degradation • Once the lungs has been cleaned, antitrypsin deactivates elastase
Role of -1 antitrypsin in elastin degradation • Without the antitrypsin enzyme, the elastase continues to consume anything in its path, including healthy lung tissue
Role of -1 antitrypsin in elastin degradation • As the healthy lung (sac) tissue is destroyed, the once stretchy walls of the sac become stiff and enlarged with air. • The enlarged sacs no longer have the ability to exchange oxygen and carbon dioxide with the bloodstream. • The lungs poor elasticity causes the lungs not to deflate properly trapping air leading to over-inflation of the lungs. • This is known as emphysema.
- KERATIN • Proteins that forms tough fibers • Found in the hair, nails, and outer epidermal layers of mammals • Constituents of intermediate filaments of cytoskeleton in certain cells • Rich in Cys, -S-S- cross-links between adjacent polypeptide chains resulting to fibers that are insoluble and resistant to stretching
GLOBULAR HEMEPROTEINS • Group of specialized protein that contains heme • Maintain a supply of oxygen essential for oxidative metabolism • MYOGLOBIN • HEMOGLOBIN
HEME • Fe+2- protoporphyrin IX • A cyclic tetrapyrrole • A planar network of conjugate double bonds absorbs visible light and colors heme deep RED.
MYOGLOBIN (Mb) • A monomeric protein (153 amino acid residues) of the red muscles • Used in some tissues, notably muscle, • as a storage reserve of O2 and • for intracellular transport of O2
Myoglobin Structure • 78% helical (the other 22% in turns and short loops, no sheet at all) • 8 - helices designated by letters A-H, in an N to C terminal direction, and connections between helices are referred to as "AB", "CD", etc. • Polar amino acid residues are found at the surface • Non polar amino acid residues (L, V, F, & M) are found at the interior except His E7 and HisF8
Myoglobin structure • The heme of myoglobin lies in crevices of helices E and F Distal histidine Proximal histidine
HEMOGLOBIN • Found exclusively in red blood cells • Its main function is to transport oxygen from the lungs to the capillaries of the tissues • The oxygen-binding properties of hemoglobin are regulated by allosteric effectors
Hemoglobin • Heterotetramer: 2 -subunits (gray and light blue) + 2 - subunits (pink and dark blue) (NOTE: designation of individual polypeptide chains with greek letters has nothing whatever to do with their secondary structural elements!)
Subunit composition of principal hemoglobin • 22 - Hemoglobin A (HgA) is the major hemoglobin in adults • 22 - Fetal hemoglobin (HbF) • 2S2 - Sickle cell hemoglobin (HbS) • 22 - Minor adult hemoglobin (HbA2) • Myoglobin and the polypeptide of hemoglobin A have almost identical secondary and tertiary structures
Quaternary structure of hemoglobin • Hgb tetramer is composed of two identical dimers, ( )1 and ()2, dimers 1 and 2 respectively • Dimers are held together by hydrophobic interactions • Ionic and H-bonding also occur between the members of the dimer
Conformational Changes resulting from oxygenation and deoxygenation of hemoglobin T form or “Taut” (tense) form • Deoxy form of Hgb • Two dimers interact through a network of ionic bonds and hydrogen bonds • Low oxygen-affinity form of hemoglobin
Conformational Changes resulting from oxygenation and deoxygenation of hemoglobin R form or Relaxed form • Binding of oxygen to Hgb (oxyhemoglobin) • Some ionic bonds and H-bonds between the dimers are ruptured thus, the polypeptide chains have more freedom of movement • High oxygen-affinity form of hemoglobin
Fe lies out of plane of heme ring Fe moves into plane of heme ring Shift from deoxy to oxy conformation
OXYGEN BINDING TO MYOGLOBIN • Can bind only one molecule of oxygen (one heme group only) • The oxygen dissociation curve for Mb is a hyperbolic shape • Mb reversibly binds a single molecule of oxygen • Mb + O2 MbO2
OXYGEN BINDING TO MYOGLOBIN HEMOGLOBIN • 4 O2 binding sites per molecule • The oxygen dissociation curve for Hgb is sigmoidal in shape • Subunits cooperate in binding oxygen
Cooperative binding of Oxygen • It means that the binding of an oxygen molecule at one heme group increases the oxygen affinity of the remaining heme group in the same Hgb molecule • This is referred to as heme-heme interaction
Cooperative Interactions (allosteric interactions) • [Greek: "allos"="other", "stereos" = space] occur when binding of one ligand at a specific site is influenced by binding of another ligand, which is called an "allostericeffector" or "modulator" • The second site is also called an allosteric site
Cooperative Interactions (allosteric interactions) • Homotropic interaction/effect • the interacting sites all bind the same ligand (e.g., binding of O2 at one site on Hb influences the binding affinity for O2 of another site) • Heterotropic interaction/effect • the interacting sitesbind different ligands
Homotropic Interaction/Effect • Positive homotropic effect • the homotropic effector increases the binding affinity for the same kind of ligand at other sites • O2 in the hemoglobin system increases the O2 binding affinity of other sites
Heterotropic Interaction/Effect • Negative heterotropic effector or allosteric inhibitor • the effector decreases the binding affinity for the primary ligand • protons, or CO2, or 2,3-BPG; all are negative heterotropic effectors of O2 binding to hemoglobin • Positive heterotropic effector, or allosteric activator • effector increases the binding affinity for the primary ligand
The Bohr Effect - effect of binding of protons (H+) and CO2 on O2 binding affinity of Hb HbO2 + H+ HbH+ + O2 Deoxyhemoglobin Increase protons or a lower pO2 Oxyhemoglobin Decrease protons or an increase pO2
Binding of Carbon dioxide • CO2 is a negative heterotropic effector (allosteric inhibitor) of O2 binding to Hb • Presence of CO2 in the tissues reduces affinity of Hb for O2 (favors deoxy, T state) in two ways: 1. CO2 lowers the pH (Bohr effect) 2. CO2 participates in formation of carbamates by the N-terminal a-amino groups of Hb: • Formation of carbamate releases H+, which contributes to the Bohr effect • Carbamate formation (CO2 binding) favors the deoxy state.
Binding of CO • CO binds tightly (but reversibly) to the Hb iron, forming carbon monoxyhemoglobin, HbCO • CO binding to one or more of the 4 heme sites of Hb shifts to relaxed conformation • The affinity of Hb for CO is 220X greater than for oxygen