Download
1 / 1
10 likes | 139 Views
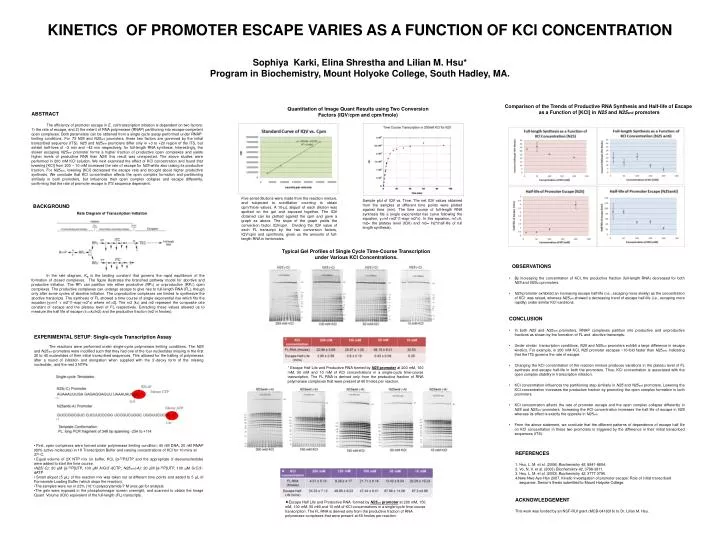
KINETICS OF PROMOTER ESCAPE VARIES AS A FUNCTION OF KCl CONCENTRATION Sophiya Karki, Elina Shrestha and Lilian M. Hsu* Program in Biochemistry, Mount Holyoke College, South Hadley, MA. Comparison of the Trends of Productive RNA Synthesis and Half-life of Escape

E N D
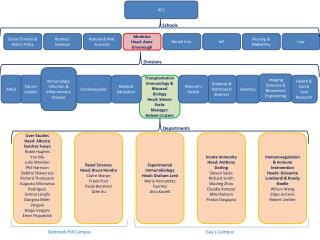
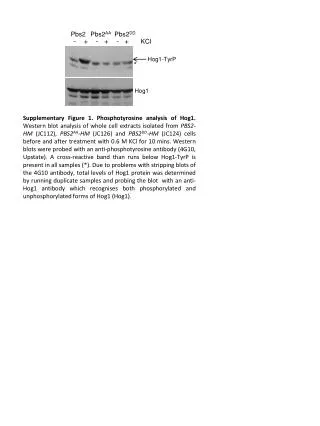
KINETICS OF PROMOTER ESCAPE VARIES AS A FUNCTION OF KCl CONCENTRATION Sophiya Karki, Elina Shrestha and Lilian M. Hsu* Program in Biochemistry, Mount Holyoke College, South Hadley, MA. Comparison of the Trends of Productive RNA Synthesis and Half-life of Escape as a Function of [KCl] in N25 and N25anti promoters Quantitation of Image Quant Results using Two Conversion Factors (IQV/cpm and cpm/fmole) ABSTRACT The efficiency of promoter escape in E. coli transcription initiation is dependent on two factors: 1) the rate of escape, and 2) the extent of RNA polymerase (RNAP) partitioning into escape-competent open complexes. Both parameters can be obtained from a single cycle assay performed under RNAP-limiting conditions. For T5 N25 and N25antipromoters, these two factors are governed by the initial transcribed sequence (ITS). N25 and N25antipromoters differ only in +3 to +20 region of the ITS, but exhibit half-lives of ~3 min and ~42 min respectively, for full-length RNA synthesis. Interestingly, the slower escaping N25antipromoter forms a higher fraction of productive open complexes and yields higher levels of productive RNA than N25; this result was unexpected. The above studies were performed in 200 mM KCl solution. We next examined the effect of KCl concentration and found that lowering [KCl] from 200 − 10 mM increased the rate of escape for N25 while also raising its productive fraction. For N25anti, lowering [KCl] decreased the escape rate and brought about higher productive synthesis. We conclude that KCl concentration affects the open complex formation and partitioning similarly in both promoters, but influences their open complex collapse and escape differently, confirming that the rate of promoter escape is ITS sequence dependent. Five serial dilutions were made from the reaction mixture, and subjected to scintillation counting to obtain cpm/fmole values. A 10-µL aliquot of each dilution was spotted on the gel and exposed together. The IQV obtained can be plotted against the cpm and gave a graph as above. The slope of the graph yields the conversion factor IQV/cpm. Dividing the IQV value of each FL transcript by the two conversion factors, IQV/cpm and cpm/fmole, gives us the amounts of full-length RNA in fentomoles. Sample plot of IQV vs. Time. The net IQV values obtained from the samples at different time points were plotted against time (min). The time course of full-length RNA synthesis fits a single exponential rise curve following the equation, y=m1+m2*(1-exp(-m3*x). In this equation, m1=0, m2= the plateau level (IQV) and m3= ln2*(half-life of full length synthesis). BACKGROUND Rate Diagram of Transcription Initiation Typical Gel Profiles of Single Cycle Time-Course Transcription under Various KCl Concentrations. OBSERVATIONS In the rate diagram, KB is the binding constant that governs the rapid equilibrium of the formation of closed complexes. The figure illustrates the branched pathway model for abortive and productive initiation. The RPc can partition into either productive (RPo) or unproductive (RPo’) open complexes. The productive complexes can undergo escape to give rise to full-length RNA (FL), though only after some cycles of abortive initiation. The unproductive complexes are limited to synthesize the abortive transcripts. The synthesis of FL showed a time course of single exponential rise which fits the equation [y=m1 + m2*(1-exp(-m3*x) where m1=0]. The m3 (kE) and m2 represent the composite rate constant of escape and the plateau level of FL respectively. Extracting these values allowed us to measure the half life of escape (t1/2=kE/ln2) and the productive fraction (m2 in fmoles). • By increasing the concentration of KCl, the productive fraction (full-length RNA) decreased for both N25 and N25antipromoters. • N25 promoter exhibited an increasing escape half-life (i.e., escaping more slowly) as the concentration of KCl was raised, whereas N25anti showed a decreasing trend of escape half-life (i.e., escaping more rapidly) under similar KCl variations. • CONCLUSION • In both N25 and N25anti promoters, RNAP complexes partition into productive and unproductive fractions as shown by the formation of FL and abortive transcripts. • Under similar transcription conditions, N25 and N25anti promoters exhibit a large difference in escape kinetics. For example, in 200 mM KCl, N25 promoter escapes ~10-fold faster than N25anti, indicating that the ITS governs the rate of escape. • Changing the KCl concentration of the reaction mixture produces variations in the plateau level of FL synthesis and escape half-life in both the promoters. Thus, KCl concentration is associated with the open complex stability in transcription initiation. • KCl concentration influences the partitioning step similarly in N25 and N25anti promoters. Lowering the KCl concentration increases the productive fraction by promoting the open complex formation in both promoters. • KCl concentration affects the rate of promoter escape and the open complex collapse differently in N25 and N25anti promoters. Increasing the KCl concentration increases the half life of escape in N25 whereas its effect is exactly the opposite in N25anti. • From the above statement, we conclude that the different patterns of dependence of escape half life on KCl concentration in these two promoters is triggered by the difference in their initial transcribed sequences (ITS). • EXPERIMENTAL SETUP: Single-cycle Transcription Assay • The reactions were performed under single-cycle polymerase limiting conditions. The N25 and N25antipromoters were modified such that they had one of the four nucleotides missing in the first 30 to 40 nucleotides of their initial transcribed sequences. This allowed for the halting of polymerase after a round of initiation and elongation when supplied with the 3’-deoxy form of the missing nucleotide, and the rest 3 NTPs. • First, open complexes were formed under polymerase limiting condition: 40 nM DNA, 20 nM RNAP (60% active molecules) in 1X Transcription Buffer and varying concentrations of KCl for 10 mins at • 37o C. • Equal volume of 2X NTP mix (in buffer, KCl, [α-32P]UTP and the appropriate 3’-deoxynucleotide) were added to start the time course. • N25(-C): 20 µM [α-32P]UTP, 100 µM A/G/3’-dCTP; N25anti(-A): 20 µM [α-32P]UTP, 100 µM G/C/3’-dATP • Small aliquot (5 µL) of the reaction mix was taken out at different time points and added to 5 µL of Formamide Loading Buffer (which stops the reaction). • The samples were run in 23% (10:1) polyacrylamide/7 M urea gel for analysis. • The gels were exposed in the phosphorimager screen overnight, and scanned to obtain the Image Quant Volume (IQV) equivalent of the full-length (FL) transcripts. * Escape Half Life and Productive RNA formed by N25 promoter at 200 mM, 100 mM, 50 mM and 10 mM of KCl concentrations in a single-cycle time-course transcription. The FL RNA is derived only from the productive fraction of RNA polymerase complexes that were present at 60 fmoles per reaction. REFERENCES 1. Hsu, L. M. et al. (2006) Biochemistry 45, 8841-8854. 2. Vo, N. V. et al. (2003) Biochemistry 42, 3798-3811. 3. Hsu, L. M. et al. (2003) Biochemistry 42, 3777-3786. 4.Nwe-Nwe Aye-Han 2007. Kinetic investigation of promoter escape: Role of initial transcribed sequence. Senior’s thesis submitted to Mount Holyoke College. ACKNOWLEDGEMENT This work was funded by an NSF-RUI grant (MCB-0418316) to Dr. Lilian M. Hsu. Escape Half Life and Productive RNA formed by N25antipromoter at 200 mM, 150 mM, 100 mM, 50 mM and 10 mM of KCl concentrations in a single-cycle time-course transcription. The FL RNA is derived only from the productive fraction of RNA polymerase complexes that were present at 60 fmoles per reaction.