Download
1 / 18
180 likes | 255 Views
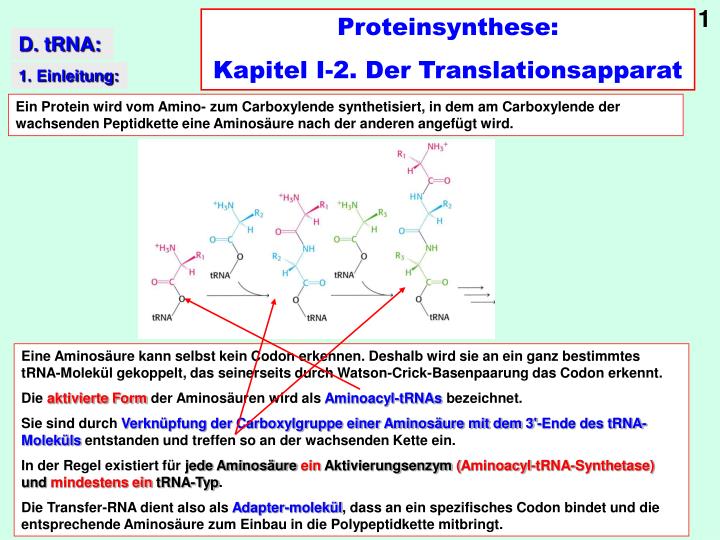
1. Proteinsynthese: Kapitel I-2. Der Translationsapparat. D. tRNA:. 1. Einleitung:. Ein Protein wird vom Amino- zum Carboxylende synthetisiert, in dem am Carboxylende der wachsenden Peptidkette eine Aminosäure nach der anderen angefügt wird.

E N D
1 Proteinsynthese: Kapitel I-2. Der Translationsapparat D. tRNA: 1. Einleitung: Ein Protein wird vom Amino- zum Carboxylende synthetisiert, in dem am Carboxylende der wachsenden Peptidkette eine Aminosäure nach der anderen angefügt wird. Eine Aminosäure kann selbst kein Codon erkennen. Deshalb wird sie an ein ganz bestimmtes tRNA-Molekül gekoppelt, das seinerseits durch Watson-Crick-Basenpaarung das Codon erkennt. Die aktivierte Form der Aminosäuren wird als Aminoacyl-tRNAs bezeichnet. Sie sind durch Verknüpfung der Carboxylgruppe einer Aminosäure mit dem 3'-Ende des tRNA-Moleküls entstanden und treffen so an der wachsenden Kette ein. In der Regel existiert für jede Aminosäure ein Aktivierungsenzym (Aminoacyl-tRNA-Synthetase)und mindestens ein tRNA-Typ. Die Transfer-RNA dient also als Adapter-molekül, dass an ein spezifisches Codon bindet und die entsprechende Aminosäure zum Einbau in die Polypeptidkette mitbringt.
2 D. tRNA: • Alle heute bekannten Molekule der Transfer-RNA haben folgende Eigenschaften: • Das Molekul ist eine einzelne Kette, die zwischen 73 und 94 Ribonucleotidenenthält (circa 25 kd). • Alle tRNAs enthalten zwischen 7 und 15 ungewöhnliche Basen (siehe Dia ). • Ungefähr die Hälfte der Nucleotide in einer tRNA sind durch Basenpaarungen zu Doppelhelices verbunden (Dia ). • Fünf Basengruppen bilden jedoch keine solchen Paare: • die Region am 3'-CCA-Ende die zum so genanten Akzeptorstamm gehört, • die T¥C-Schleife, die ihren Namen der Sequenz Ribothymin-Pseudouracil-Cytosin verdankt, • der ,,Extraarm", der eine variable Anzahl von Resten enthält, • die DHU-Schleife mit mehreren Dihydrouracilbausteinen und die Anticodonschleife. • Wegen der Strukturvielfalt, die durch diese Kombination aus Helices und Schleifen mit modifizierten Basen entsteht, sind alle tRNA strotz ihrer ähnlichen Gesamtstrukturen einzeln zu unterscheiden. • 4. Das 5'- Ende der tRNA ist phosphoryliert (pG). • 5. Die aktivierte Aminosäure wird an eine Hydroxylgruppe des Adenosinrestes angeheftet, der sich am Ende des 3'-CCA-Bestandteils des Akzeptorstammes befindet. Dieser Abschnitt am 3'-Ende ist bei reifen tRNAs einzelsträngig. • 6. Das Anticodon befindet sich in einer Schleife ungefähr in der Mitte der Sequenz. 2. Das gemeinsame Konstruktionsprinzip: Die Matrizenerkennungsstelle auf der tRNA ist eine Sequenz von drei Basen, die man als Anticodon bezeichnet. Das Anticodon erkennt auf der mRNA jeweils eine komplementäre Sequenz von drei Basen, das Codon.
3 D. tRNA: 3. Die dreidimensionale Struktur: • Die dreidimensionale Struktur eines tRNA-Moleküls hat folgende wichtige Eigenschaften: • 1. Das Molekul ist L-förmig. • 2. Es gibt zwei scheinbar ununterbrochene Doppelhelixabschnitte: • Die Helix mit dem 5'- und 3'-Ende liegt oberhalb der Helix, die in der T¥C-Schleife endet, sodass sie einen Arm des L bilden. • Die beiden übrigen Helices (DHU- und Anticodon Schleifen) lagern sich zu dem anderen Arm zusammen. • 3. In den Abschnitten, die keine Helix bilden, sind die meisten Basen an Wasserstoffbrücken beteiligt, die allerdings nicht denen in Watson-Crick-Basenpaaren entsprechen. • 4. Das CCA-Ende mit der Aminosäureanhechtungsstelle ragt an einem Ende aus dem L heraus. Dieser einzelsträngige Abschnitt kann seine Konformation während der Aktivierung der Aminosäure und der Proteinsynthese ändern. • 5. Die Anticodonschleife befindet sich am anderen Ende des L, weit (7,8 nm entfernt von der Anheftungsstelle für die Aminosäuren), sodass die drei Basen des Anticodons gut zugänglich sind. • Mit seinem Aufbau eignet sich das tRNA-Molekül also gut für seine Adapter-funktion: Das Anticodon ist zugänglich und kann mit einem geeigneten Codon auf der mRNA in Wechselwirkung treten, und das Ende, das mit der aktivierten Aminosäure gekoppelt ist, liegt an der richtigen Stelle für die Ausbildung der Peptidbindung.
4 D. tRNA: 4. Das Prozessieren von Vorläufer RNA-Ketten: Bestimmte Transkripte enthalten Aneinanderreihungen verschiedener Arten von tRNAs oder mehrerer Kopienderselben tRNA die auseindergespleisst werden. Reife tRNAs werden durch Spaltung und Modifikation von Vorläufer RNA-Ketten erzeugt. Die Ribonuclease P erzeugt in E. coli das korrekte 5'-Ende aller tRNA-Moleküle. Dieses interessante Enzym enthält ein katalytisch aktives RNA-Molekül(Ribozym). Ein zweiter Prozessierungstyp ist das Anfügen von CCA an das 3'-Ende von tRNA-Molekülen, die diese terminale Sequenz noch nicht besitzen. Dies gescheht in zwei Schritten. Ein dritter Prozessierungstyp ist das herausspleißen eines Introns das 3’ vom Anticodon (also stromabwärts) positionniert ist (Vorgang wird nicht gezeigt, gescheht in drei Schritten). Der letzter Schritt der Prozessierung ist die Modifikation von bestimmten Basen und Riboseeinheiten.
5 D. tRNA: 5. Der genetischer Code: Eigenschaften. • Eigenschaften des genetischen Codes: • 1. Drei Nucleotide codieren eine Aminosäure. • 2. Der Code könnte nichtüberlappend oder überlappend sein: • Betrachten wir die Sequenz ABCDEF: • Bei einem überlappenden Code stünde ABC für die erste Aminosäure, BCD für die zweite und CDE für die dritte und so weiter. • Bei einem nichtüberlappenden Code spezifiziert ABC die erste, DEF die zweite und so weiter. • Der Code ist nichtüberlappend • Der Code verfügt über keine Zeichensetzung. Im Prinzip könnte eine Base (hier Q (Rot) genannt) als ,,Komma" zwischen den Basentripletts dienen: ... QABCQDEFQGHIQJKLQ... • Die Basensequenz wird vielmehr fortlaufend von einem bestimmten Startpunkt an abgelesen. Eine Zeichensetzung gibt es nicht. • Der genetische Code ist degeneriert. Manche Aminosäuren werden von mehr als einem Codon codiert, denn es gibt 64 mögliche Tripletts und nur 20 Aminosäuren. • Tatsächlich stehen 61 der 64 möglichen Tripletts für eine bestimmte Aminosäure, die drei übrigen signalisieren das Ende der Translation und werden daher als Stoppcodons bezeichnet. Für die meisten Aminosäuren gibt es daher mehr als ein Codewort.
6 D. tRNA: 6. Die degenerierte Natur des genetische Codes: Da der Code in hohem Maße degeneriert ist, werden lediglich Tryptophan und Methionin von einem einzigen Triplett codiert. Für die übrigen 18 Aminosäuren gibt es zwei oder mehr Codons. Leucin, Arginin und Serin werden sogar von jeweils sechs Codons spezifiziert. Die Zahl der Codons für eine bestimmte Aminosäure korreliert mit der Häufigkeit ihres Auftretens in Proteinen. Codons, welche die gleiche Aminosäure codieren, heißen Synonyme. CAU und CAC sind zum Beispiel Synonyme für Histidin. Synonyme verteilen sich nicht rein zufällig über die Tabelle des genetischen Codes. Eine Aminosäure, die von zwei oder mehreren Synonymen codiert wird, taucht nur in einem einzigen Kästchen auf (sofern es nicht mehr als vier Synonyme gibt). Die Aminosäuren in einem Kästchen werden von Codons spezifiziert, die in den ersten beiden Basen übereinstimmen, sich aber in der dritten unterscheiden, wie etwa GUU, GUC, GUA und GUG verdeutlichen. Somit unterscheiden sich die meisten Synonyme nur in der letzten Base des Tripletts. Anmerkung: Wenn man die Stellung der Basen in einem Codon kennt, kann man die entsprechende Aminosäure finden. Zum Beispiel spezifiziert das Codon 5'-AUG-3' auf der mRNA Methionin und CAU Histidin. UAA, UAG und USA sind Terminationssignale. AUG ist Teil des Initationssignals, zusätzlich zu seiner Rolle als Codewort für Methionin innerhalb von Ketten.
7 D. tRNA: 8. Der genetischer Code : 7. Die degenerierte Natur des genetische Codes: Bedeutung. Welche biologische Bedeutung bringt die massive Degeneriertheit des genetischen Codes mit sich? Wäre der Code nicht degeneriert, würden 20 Codons Aminosäuren codieren und 44 eine Kettentermination bewirken. So wäre die Wahrscheinlichkeit für Mutationen, die zum Kettenabbruch führen, weitaus größer. Durch einen Kettenabbruch entstehen im Allgemeinen inaktive Proteine, während der Ersatz einer Aminosäure durch eine andere meistens relativ harmlos ist. Somit minimiert die Degeneriertheit die schädlichen Auswirkungen von Mutationen. Des Weiteren ist vermutlich von Belang, dass die Basenzusammensetzung aufgrund der Degeneriertheit des genetischen Codes über einen weiten Bereich variieren kann, ohne dass es dadurch zu einer veränderten Aminosäuresequenz der von der DNA codierten Proteine kommt. Der GC- Gehalt von Bakterien-DNA reicht von weniger als 30 bis zu über 70. DNA- Moleküle mit ganz verschiedenem GC-Gehalt können trotzdem die gleichen Proteine codieren, wenn durchweg verschiedene Synonyme verwendet würden.
8 D. tRNA: 8. Verschiedene tRNA Typen: • Initiator-tRNA: • Wie wir später sehen werden, ist bei Prokaryoten die erste Aminosäure N-Formyl-methionin, und bei Eukaryoten Methionin. Das Initiationscodon der mRNA lautet entsprechend AUG, aber an der Initiation ist eine besondere tRNA beteiligt. Diese heißt in ihrer Aminoacylform i-tRNAMet, oder i-tRNAMet-f (der Index ,,i" steht für Initiation", das ,,f" weist darauf hin, dass die Aminosäure in vitro formyliert werden kann). Es weist ein Besonderheit auf indem die 5’Base keine Basenpaarung angeht. • Elongator-tRNA: • Diese Elongator-tRNA (abgekürzt tRNAn) lehst AUG Codons im Inneren der Proteinkette und baut Methionin ein. Für die Verknüpfung des Methionins mit den zwei verschiedenen tRNAs ist dieselbe Aminoacyl-tRNA-Synthetase verantwortlich. • 3. Iso-empfänger-tRNA-Moleküle: siehe Dia 8 und 9
9 D. tRNA: 9. : durch die Ungenauigkeit der Baasenpaarung « Wobble » erkennen manche tRNA-Moleküle mehrere Codons: Codon und Anticodon sind anti-parallel angeordnet: die erste Base des Anticodons bindet an der dritte Base des Codons. In dem Diagramm kennzeichnet das „’“ jeweils die komplementären Base. Nach diesem Modell kann jedes Anticodon nur ein ganz bestimmtes Codon erkennen. In Wirklichkeit erkennen manche reinen tRNA-Moleküle mehrere Codons: 8 Aminosäuren werden von 4 Codons spezifiert die nur in der 3. Base variieren. Die Wobble Hypothese von Francis Crick sagt voraus dass die sterischen Kriterien für die Paarung der dritten Base des Codons mit der 1. Base des Anticodons weniger streng sein könnten als für die beiden anderen. Mit Einbeziehung des Inosins, dass in mehreren Anticodons vorkommt, ergibt sich für die Paarung der dritten Base eines Codons eine gewisse sterische Freiheit (,,Wobble"). Volgende Kombinationen sind möglich.
10 D. tRNA: 9. : durch das « Wobble » der Baasenpaarung erkennen manche tRNA-Moleküle mehreren Codons: Iso-Empfänger. Die Wobble-Hypothese ist heute allgemein anerkannt: die Anticodons von tRNA-Molekülen mit bekannter Sequenz binden an die Codons, die aufgrund dieser Hypothese vorausgesagt werden. Allgemeine Aussagen über die Wechselwirkungen zwischen Codon und Anticodon : 1. Die beiden ersten Basen eines Codons gehen die üblichen Paarungen ein. Ihre Erkennung erfolgt sehr präzise. Codons, die sich in einer der beiden ersten Basen unterscheiden, müssen also von unterschiedlichen tRNAs erkannt werden. Zum Beispiel codieren sowohl UUA als auch CUA Leucin, sie werden aber von unterschiedlichen tRNAs abgelesen. Diese tRNA-Molekülen werden Iso-empfäger genannt. 2. Die erste Base eines Anticodons bestimmt darüber, ob ein bestimmtes tRNA-Molekül ein, zwei oder drei Codons erkennt: C oder A (ein Codon), U oder G (zwei Codons), oder I (drei Codons) (Siehe Dia 9). Dass der genetische Code degeneriert ist, ergibt sich also zum Teil aus der Ungenauigkeit (Wobble) in der Paarung der dritten Base des Codons mit der ersten Base des Anticodons. Hier liegt ein Grund dafür, dass das ungewöhnliche Nucleosid Inosin in Anti-codons so häufig vorkommt. Inosin schafft die Möglichkeit, dass ein bestimmtes tRNA-Molekül die maximale Zahl von Codons erkennen kann. Warum wird eine Wobble-Basenpaarung in der dritten Position des Codons zugelassen, nicht aber in den beiden ersten? Die 30S-Untereinheit enthält zwei Adeninbasen (A1492 und A1498 in der 16S-RNA), die auf der Seite der kleinen Furche Wasserstoffbrücken mit dem Komplex aus Codon und Anticodon ausbilden. Mittels dieser Wechselwirkungen wird überprüft, ob in den beiden ersten Positionen des Codon-Anticodon-Doppelstranges Watson-Crick-Basenpaare vorhanden sind. An der dritten Position steht kein derartiges Überprüfungshilfsmittel zur Verfügung, sodass dort vielgestaltigere Basenpaare zugelassen werden. Das Ribosom ist also an der Decodierung der Wechselwirkungen zwischen Codon und Anticodon aktiv beteiligt.
11 D. tRNA: 10. Der genetische Code ist nahezu universell : Ist der genetische Code bei allen Organismen derselbe? Die Basensequenzen vieler Gene von Wildtypen und Mutanten sind ebenso bekannt wie die Aminosäuresequenzen der von ihnen codierten Proteine. In allen Fällen entsprechen der Nucleotidwechsel im Gen und der Aminosäurewechsel in dem Protein den Vorgaben des genetischen Codes. Weiterhin können mRNAs durch die Proteinsyntheseapparate ganz verschiedener Spezies richtig übersetzt werden. Dies spricht sehr dafür, dass der genetische Code universell ist. Der genetische Code ist « de Facto » fast vollständig universell. Abweichungen treten nur in Mitochondrien auf sowie bei Arten - etwa den Ciliaten -, die in der Evolution der Eukaryoten frühzeitig einen eigenen Weg beschritten haben. Warum ist der Code über Jahrmilliarden der Evolution vom Bakterium zum Menschen nahezu unverändert geblieben? Eine Mutation, die das Ablesen der mRNA verändert, würde die Aminosäuresequenzen der meisten - vielleicht sogar aller - Proteine abwandeln, die von dem betreffenden Organismus erzeugt werden. Viele solcher Änderungen wären ohne Zweifel tödlich und so würden Mutationen mit derart weitreichenden Konsequenzen durch die natürliche Selektion sofort ausgemerzt.
12 E. Das Ribosom: a. Komponenten: 1. Das prokaryotische Ribosom: • Ribosomen sind Molekülmaschinen, die das Wechselspiel zwischen beladenen tRNAs, mRNA und Proteinen koordinieren und so die Proteinsynthese ermöglichen. • Ein Ribosom ist eine Ribonucleoproteinanordnung mit einer Masse von rund 2700 kd, einem Durchmesser von ungefähr 20 nm und einem Sedimentationskoeffizienten von 70S. • Die 20000 Ribosomen einer Bakterienzelle machen nahezu ein Viertel ihrer Masse aus. • Ein Ribosom (70S) enthält eine kleine (30S) und eine große (50S) Untereinheit. • Die 30S-Untereinheit enthält ein RNA-Molekül von 16S und 21 verschiedene Proteine (S1 bis S21). • Die 50S-Untereinheit enthält zwei RNA-Moleküle, (23S und 5S) und 34 verschiedene Proteine (L1 bis L34). • Ein Ribosom enthält: • ein Exemplar jedes RNA-Moleküls, jeweils zwei Molekule der Proteine L7 und L12 sowie jeweils ein Molekul der anderen Proteine. • Das Protein L7 ist mit L12 identisch, nur mit dem Unterschied, dass sein Aminoende acetyliert ist. • Nur ein einziges Protein kommt in beiden Untereinheiten vor: S20 ist identisch mit L26, und befindet sich an der Wechselwirkungsebene zwischen beide Untereinheiten. 4
13 C. Das Ribosom: b. In vitro rekonstitution: 1. Das prokaryotische Ribosom: Sowohl die 30S- als auch die 50S-Untereinheit kann man in vitro aus ihren Protein- und RNA-Bausteinen zusammensetzen. Vorausetzung sind die richtigen Konditionen (pH, ionische Konzentrationen, Temperatur). Jedes Protein bindet sich an der jeweilige rRNA nach einer strikt festgelegte Reihenordnung. Diese Rekonstitution ist ein bemerkenswertes Beispiel für das Prinzip, dass sich supramolekulare Komplexe spontan aus ihren makromolekularen Bausteinen bilden können. Auf Zellebene, startet die Zusammensetzung der 30S Untereinheit sogar schon bevor die rRNA Transkription zu Ende geht. Das 5-Ende hat in der Tat eine hohe Affinität für bestimmte der Proteine.
14 C. Das Ribosom: c. Das ribosomale Operon: 1. Das prokaryotische Ribosom: Im Gegensatz zu den mRNAs werden rRNAs durch Spaltung einer Prä-RNA-Kette erzeugt. Drei verschiedene Arten von rRNAs und eine tRNA werden aus einem einzigen RNA-Primärtranskript, das auch Spacer-Regionen enthält, herausgeschnitten. Die Ribonuclease III, die diesen Vorläufer der rRNA spaltet, schneidet 5S-, 16S- und 23S-rRNA-Vorläufer aus dem Primärtranskript, indem sie doppelhelikale Stamm-Schleife- Strukturen an spezifischen Stellen spaltet. Einige Basen der rRNA werden methyliert.
15 C. Das Ribosom: d. Die dreidimensionale Struktur 1: 1. Das prokaryotische Ribosom: Heute kennt man durch die Methoden der Röntgenstrukturanalyse die Struktur des vollständigen 70S-Ribosoms in fast allen Bereichen bis zur Ebene einzelner Atome. Die drei in dem Ribosom enthaltenen RNAs von 5S, 16S und 23S sind für Aufbau und Funktion der Ribosomen von entscheidender Bedeutung. Jahrelang glaubte man, die Ribosomenproteine seien für die Koordination der Proteinsynthese verantwortlich; die ribosomale RNA hielt man vorwiegend für ein Strukturgerüst. Nach heutiger Kenntnis steht fest dass die entscheidenden Stellen in den Ribosomen fast ausschließlich aus RNA bestehen. Die Proteine leisten dazu nur einen bescheidenen Beitrag und es ist wahrscheinlich dass die Ribosomen ursprünglich ausschließlich aus RNA bestanden und dass die Proteine erst später zur Feinabstimmung ihrer Funktionseigenschaften hinzukamen.
16 C. Das Ribosom: e. Die dreidimensionale Struktur 2: 1. Das prokaryotische Ribosom: Die ribosomalen RNAs (wie hier das 16S-rRNA) sind zu einer definierten Struktur mit vielen kurzen Doppelstrangabschnitten gefaltet. Die durch Röntgenanalyse ermittelte tertiärstruktur des 16S-rRNA.
16 C. Das Ribosom: f. Die Funktionsmerkmale: 1. Das prokaryotische Ribosom: Das 3’-Ende der 16S-rRNA Bindet die mRNA (Kapitel 2). Anticodons der tRNAs binden an einer Spalte der kleine Untereinheit (Kapitel 2). 4 Kopieen des L7-L12 Komplexes verleihen eine GTPase Aktivität (Kapitel 2). tRNAs binden das Ribosom in zwei Stelllen, A und P site genannt. Das Peptidyl-Transferase dass die Aminosäuren zum Zweck des Peptiverbindungs Reaktion positioniert ist enthalten in der großen Untereinheit, und bindet hier an der 23S-rRNA. Die wachsende Polypeptidkette (blau) tretet ans Licht aus einem Tunnel innerhalb der grossen Untereinheit.
17 C. Das Ribosom: 2. Das eukaryotische Ribosom: Ein eukarytisches Ribosom ist größer (80S) als sein prokarytosischer Kollege (70S). Es enthält eine kleine (40S) und eine große (60S) Untereinheit. Die 40S-Untereinheit enthält ein RNA-Molekül von 18S und 33 verschiedene Proteine (S1 bis S33). Die 60S-Untereinheit enthält drei RNA-Moleküle, (5S, 5,5S, und 28S) und 49 verschiedene Proteine (L1 bis L49).





















