Download
1 / 37
470 likes | 959 Views
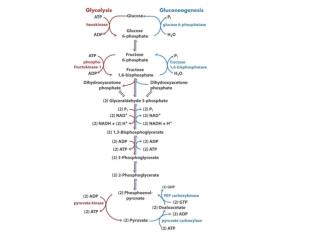
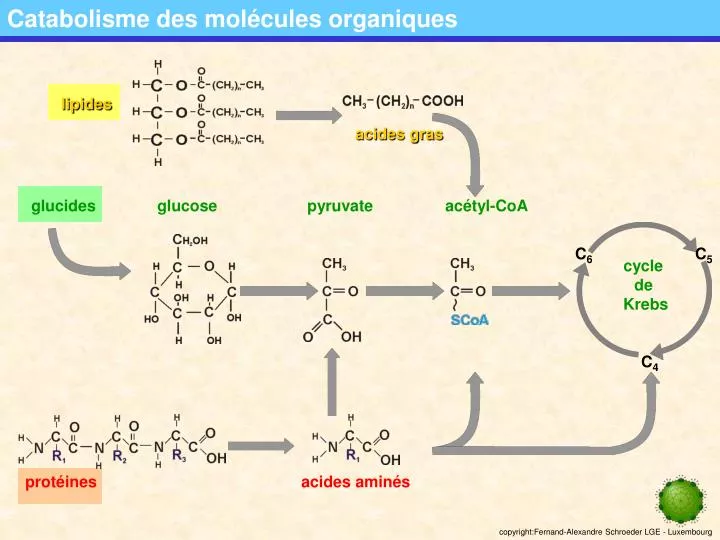
Catabolisme des molécules organiques. lipides. glucose. protéines. acides aminés. acétyl-CoA. glucides. C 6. C 5. cycle de Krebs. pyruvate. C 4. acides gras. La dégradation du glucose: la glycolyse. glucose. glucose-6-P. fructose-6-P. fructose-1;6-diP. ATP. ADP. ATP. ADP.

E N D
Catabolisme des molécules organiques lipides glucose protéines acides aminés acétyl-CoA glucides C6 C5 cycle de Krebs pyruvate C4 acides gras
La dégradation du glucose: la glycolyse glucose glucose-6-P fructose-6-P fructose-1;6-diP ATP ADP ATP ADP Bilan de cette étape: consommation de 2 molécules d’ATP: (-2ATP)
La dégradation du glucose: structure de l’ATP adénine phosphates ribose ATP ADP + EAU + Pi
La dégradation du glucose: structure du NADH + H+ ribose ribose phosphates adénine nicotinamide nicotinamide acide nicotinique (niacine ou vitamine B3 anciennement vitamine PP)
La dégradation du glucose: la glycolyse - suite fructose- 1;6-diP phosphoglycér- aldéhyde diphospho- glycérate phospho- glycérate pyruvate Pi H2O NAD+ ADP NADH+H+ ATP ADP ATP Bilan de cette étape: production de 4 molécules d’ATP: (+ 4ATP) production de 2 molécules de NADH+H+: ( + 2NADH2)
La dégradation du glucose: la glycolyse – bilan provisoire glucose pyruvate Bilan de la glycolyse: consommation de 2 molécules d’ATP: - 2 ATP production de 4 molécules d’ATP: + 4 ATP production de 2 molécules de NADH+H+: + 2 NADH2 production de 2 molécules d’ATP: 2 ATP production de 2 molécules de NADH+H+: 2 NADH2
La dégradation du glucose: décarboxylation ou oxydation du pyruvate pyruvate acétyl-CoA NAD+ NADH+H+ CO2 HS-CoA Bilan de l’oxydation du pyruvate: production de 2 molécules de NADH+H+: + 2 NADH2
La dégradation du glucose: structure de la coenzyme A SH b-mercaptoéthylamine acide pantothénique (=vitamine B5) adénine ribose 3-phosphate CoA notation: CoA-SH ou HS-CoA
La dégradation du glucose: cycle de Krebs acétyl-CoA HS-CoA NADH+H+ NAD+ citrate malate H2O H2O oxalate H2O isocitrate fumarate a-céto- glutarate NAD+ NADH+H+ succinyl- CoA FADH2 FAD NAD+ succinate NADH+H+ ATP ADP + Pi H2O HS-CoA CO2 H2O Bilan de cette étape: + 6 NADH2 + 2 FADH2 + 2 ATP CO2 HS-CoA
La dégradation du glucose: structure du FAD/ H2 forme oxydée: FAD Forme réduite: FADH2 riboflavine ( vitamine B2) ribose adénine phosphates
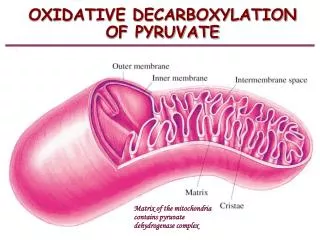
La dégradation du glucose: localisation des voies métaboliques ATP = phosporylation liée au substrat glucides cytosol mitochondrie C6 C5 cycle de Krebs C4 Le complexe enzymatique (la pyruvate déshydrogénase) responsable de l’oxydation du pyruvate est logé dans la membrane mitochondriale. ATP pyruvate glucose pyruvate acétyl-CoA ATP
La dégradation du glucose: ultrastructure de la mitochondrie membrane externe membrane interne espace inter- membranaire crête mitochondriale complexes ATP-synthétase mitoribosome ADN mitochondrial matrice
La dégradation du glucose: ultrastructure de la mitochondrie 5 5
La dégradation du glucose: phospophorylation liée au substrat On voit que la glycolyse et le cycle de Krebs produisent une petite quantité d’ATP. Cet ATP est formé grâce au transfert enzymatique direct d’un groupement phosphate à partir d’un phosphate inorganique ou d’un substrat à l’ADP. Cette phosphorylation de l’ADP s’appelle phosphorylation liée au substrat.
La dégradation du glucose: exemples de phospophorylations liées au substrat diphospho- glycérate phospho- glycérate pyruvate H2O ADP + Pi ATP HS-CoA ADP ATP ADP ATP succinyl- CoA succinate H2O lors de la glycolyse lors du cycle de Krebs
La dégradation du glucose: bilan provisoire pour la dégradation d’une molécule de glucose Bilan provisoire: glucose 6CO2 production de 10 molécules de NADH+H+: 10 NADH2 production de 2 molécules des FADH2; 2 FADH2 production de 4 molécules d’ATP: 4 ATP GLYCOLYSE Bilan de la glycolyse: production de 2 molécules de NADH+H+: + 2 NADH2 production de 2 molécules d’ATP: + 2 ATP OXYDATION DU PYRUVATE Bilan de l’oxydation du pyruvate: production de 2 molécules de NADH+H+: + 2 NADH2 CYCLE DE KREBS Bilan du cycle de Krebs: production de 6 molécules de NADH+H+: + 6 NADH2 production de 2 molécules des FADH2; + 2 FADH2 production de 2 molécules d’ATP: + 2 ATP
Chaîne respiratoire ou système de transporteurs d’électrons protéine contenant du Fe-S cytochromes coenzyme Q déshydrogénase flavoprotéique (=flavoprotéine) mitochondrie matrice chaîne de transport des électrons espace intermembranaire ATP synthétase
Chaîne respiratoire: principales oxydo-réductions et sites de couplage 2H+ H2O NAD+ ½ O22- FAD 2H+ 2H+ 2H+ 2H+ 2e- 2e- 2e- 2e- H+ H+ H+ NADH+H+ ½ O2 FADH2 2H+ H+ H+ H+ 2e- H+ H+ H+ 2e- 2e- 2e- 2e- H+ H+ H+ H+ H+ H+ Au cours du transport des électrons à l’oxygène il y a translocation de protons de la matrice vers l’espace intermembranaire. Cette translocation se réalise en trois sites et fait intervenir des protéines fer-soufre. Dans une première étape, deux protons accompagnés de deux électrons sont arrachés du NADH2 pour passer sur la flavoprotéine (Fe-S) et ensuite sur la CoQ. Il faut noter qu’à ce niveau les deux atomes d’hydrogène ne sont plus transférés mais libérés dans la matrice et les électrons passent sur les cytochromes. ( Il faut donc deux systèmes cytochromiques pour deux électons !) Finalement les électrons réduisent l’oxygène qui se combine à deux protons pour former une molécule d’eau.
Chaîne respiratoire: phosphorylation de l’ADP H+ ATP La translocation des protons de la matrice vers l’espace intermembranaire engendre un gradient électrochimique. Le retour des protons dans la matrice à travers la base hydrophobe des ATPases et leur pédoncule est responsable de la phosphorylation de l’ADP. Comme cette phosphorylation est liée à l’oxydation de l’hydrogène on parle de phosphorylation oxydative. H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ ATP-synthétase H+ ADP + Pi
Chaîne respiratoire: prinicipe de l’oxydo-réduction ATP substrat oxydée ATP ATP Site I Site II Site III FADH2 FAD H2O 2H+ ADP+ Pi ADP+ Pi ½ O22- 2 a3 (Fe3+) substrat NAD+ FP-H2 CoQ 2 b (Fe2+) 2 c (Fe3+) 2 a (Fe2+) NADH+H+ FP CoQ-H2 2 b (Fe3+) 2 c (Fe2+) 2 a (Fe3+) 2 a3 (Fe2+) ½ O2 ADP+ Pi DONC: Chaque molécule de NADH+ + H+ donne 3 molécules d’ATP Chaque molécule de FADH2 donne 2 molécules d’ATP
La dégradation du glucose: localisation complète des voies métaboliques ATP = phosporylation liée au substrat glucides ATP cytosol mitochondrie = phosporylation oxydative C6 C5 cycle de Krebs C4 ATP pyruvate glucose ATP acétyl-CoA ATP
La dégradation du glucose: les navettes mitochondriales Problème: Le NADH+H+ dans cytosol (produit lors des réactions de déshydrogénation de la glycolyse) ne peut pas franchir pas la membrane mitochondriale interne. Solution: NADH+H+ est oxydé en NAD+ pour céder l’hydrogène à des molécules “navettes” capables de traverser la membrane mitochondriale interne. Il existe deux navettes: • Navette malate / aspartate • Navette glycérol phosphate
La dégradation du glucose: la navette malate/aspartate NADH+H+ NAD+ malate déshydrogénase cytosolique et mitochondriale oxalate malate
La dégradation du glucose: la navette malate/aspartate NAD+ NADH+H+ oxalate oxalate malate malate NAD+ NADH+H+ mitochondrie Bilan: 1 molécule de NADH+H+ cytosolique donne 1 molécule de NADH+H+ mitochondriale.
La dégradation du glucose: la navette glycérol phosphate NADH+H+ NAD+ glycérophosphate déshydrogénase cytosolique phospho- glycérate phospho- dihydroxyacétone Le glycérol phosphate peut traverser la membrane mitochondriale externe.
La dégradation du glucose: la navette glycérol phosphate CoQ CoQH2 glycérophosphate déshydrogénase mitochondriale phospho- glycérate phospho- dihydroxyacétone Au niveau de la membrane mitochondriale interne il y a oxydation du glycérol phosphate en PDHA avec transfert des électrons et H+ sur CoQ. La glycérophosphate déshydrogénase mitochondriale est en effet une flavoprotéine.
La dégradation du glucose: la navette malate/aspartate phospho- glycérate phospho- dihydroxyacétone mitochondrie 2H+ + 2é Bilan: 1 molécule de NADH+H+ cytosolique donne 1 molécule de FADH2 mitochondriale.
La dégradation du glucose en aérobiose: bilan énergétique définitif Bilan provisoire: glucose 6CO2 production de 10 molécules de NADH+H+: 10 NADH2 production de 2 molécules des FADH2; 2 FADH2 production de 4 molécules d’ATP: 4 ATP Chaque molécule de NADH+ + H+ donne 3 molécules d’ATP Chaque molécule de FADH2 donne 2 molécules d’ATP Donc: glucose CO2 équivalent de 10 molécules de NADH+H+: 30 ATP équivalent de 2 molécules des FADH2; 4 ATP production de 4 molécules d’ATP: 4 ATP production totale d’ATP 38 ATP C6H12O6 + 6O2 6CO2 + 6H2O + 38 ATP Remarque: Selon la navette utilisée une molécule de glucose va générer 36 ou 38 molécules d’ATP!
Respiration anaérobie: fermentation lactique lactate 2ATP glucose 2NAD+ 2NAD+ pyruvate 2ADP + 2Pi 2NADH2 2NADH2 conséquences: suite au faible rendement énergétique, le substrat est vite consommé, le pH du milieu décroît en plus le lactate est toxique ! mode de respiration en cas d’absence d’oxygène p.ex. dans les muscles des Mammifères après exercice physique prolongé Bilan de la glycolyse en anaérobiose (fermentation lactique): production de 2 molécules d’ATP par molécule de glucose
Respiration anaérobie: fermentation alcoolique acétaldéhyde éthanol 2ATP glucose 2CO2 2NADH2 2NAD+ pyruvate 2ADP + 2Pi 2NAD+ 2NADH2 mode de respiration en cas d’absence d’oxygène réservé à quelques rares microorganismes comme la levure p.ex.. des expériences récentes ont montré l’existence de la fermentation alcooliques chez la carpe en hibernation dans les fonds de vase pauvres en oxygène. conséquence: le taux d’alcool du milieu augmente; l’éthanol est toxique ! Bilan de la glycolyse en anaérobiose (fermentation alcoolique): production de 2 molécules d’ATP par molécule de glucose
Dégradation du glucose: bilans énergétiques et rendements C6H12O6 + 6O2 6CO2 + 6H2O + 38 ATP énergie libre d’une mole de glucose: DG = -686 kcal/mol énergie libre d’une mole d’ATP: DG = -7,3 kcal/mol énergie libre de 38 moles d’ATP: DG = -7,3 * 38 = -277,4 kcal rendement de la respiration: 277,4 / 686 = 0,404 ( = 40,4%) (à titre de comparaison: rendement d’une voiture: à peu près 25 %) A peu près 60 % de l’énergie libre contenue dans une mole de glucose est dissipée sous forme de chaleur. Une partie de cette chaleur est utilisée chez les oiseaux et le mammifères pour maintenir leur température corporelle élevée; le reste se perd par rayonnement, sudation ou autres mécanismes de refroidissement. rendement de la fermentation: 14,6 / 686 = 0,021 ( = 2,1%)
Utilisation de l’ATP dans la contraction musculaire • L'énergie mécanique de la contraction musculaire provient directement de l'énergie chimique (ATP). Pendant l'activité musculaire, la régénération de l'ATP se fait suivant 3 voies successives: • par interaction de l'ADP avec la créatine phosphate: (< 10 secondes) • par respiration cellulaire anaérobie (< 1minute) • par respiration cellulaire aérobie (> 1 minute)
Utilisation de l’ATP dans la contraction musculaire 1. Au début d'une activité musculaire, l'ATP emmagasiné dans les muscles actifs est consommé en 6 secondes environ. Un système de production rapide d'ATP se met en place, en attendant que les voies métaboliques s'adaptent à la demande accrue d'ATP. L'ADP se couple alors à la créatine phosphate (créatine kinase), composé à haute énergie emmagasiné dans les muscles. Il en résulte un transfert presque instantané d'énergie et la formation d'une molécule d'ATP. Une puissance musculaire maximale peut ainsi être maintenue pendant 10 à 15 secondes (sprint sur 100 m). créatine phosphate
Utilisation de l’ATP dans la contraction musculaire 1. phosophocréatine ( < 15 secondes) créatine kinase créatine phosphate créatine ADP ATP
Utilisation de l’ATP dans la contraction musculaire 2. Respiration cellulaire anaérobie : la glycogénolyse. Pour des exercices musculaires de plus longue durée, le glycogène musculaire doit être dégradé. Les réserves de glycogène du muscle sont transformées en acide lactique via le glucose-6-phosphate, avec production de 2 molécules d'ATP par molécule de glucose (faible rendement énergétique). La glycolyse anaérobie commence plus tardivement que la dégradation de la créatine phosphate (au maximum après 30 secondes) et produit de l'ATP 2,5 fois plus vite que la voie aérobie. Ainsi, lorsqu'il faut de grandes quantités d'ATP pendant de courtes périodes d'activité musculaire soutenue (30-40 secondes), la voie anaérobie en fournit une grande partie. Ensemble, les réserves d'ATP et de créatine phosphate et le système glycogénolyse-acide lactique peuvent entretenir une activité musculaire pendant presque une minute.
Utilisation de l’ATP dans la contraction musculaire 2. glycogénolyse avec respiration anaérobie ( < 1 minute) glycogène (>100 000 a-glucose) glucose-6-P pyruvate lactate
Utilisation de l’ATP dans la contraction musculaire 3. Respiration cellulaire aérobie : la phosphorylation oxydative du glucose et des acides gras.(> 1 minute) Pour que l'effort soit maintenu, il faut absolument qu'il y soit hydrolyse aérobie du glucose et des acides gras. Lors des contractions lentes ou au repos, la plus grande partie de l'approvisionnement en ATP est assurée par la respiration cellulaire aérobie, qui utilise l'énergie fournie par la dégradation des acides gras. Lorsque les muscles se contractent de façon plus soutenue, c'est le glucose qui devient la principale source d'énergie. Globalement, l'oxydation complète d'une molécule de glucose en CO2 et en eau fournit 38 molécules d'ATP (rendement énergétique élevé). C6H12O6 + 6O2 6CO2 + 6H2O + 38 ATP