Download
1 / 12
140 likes | 469 Views
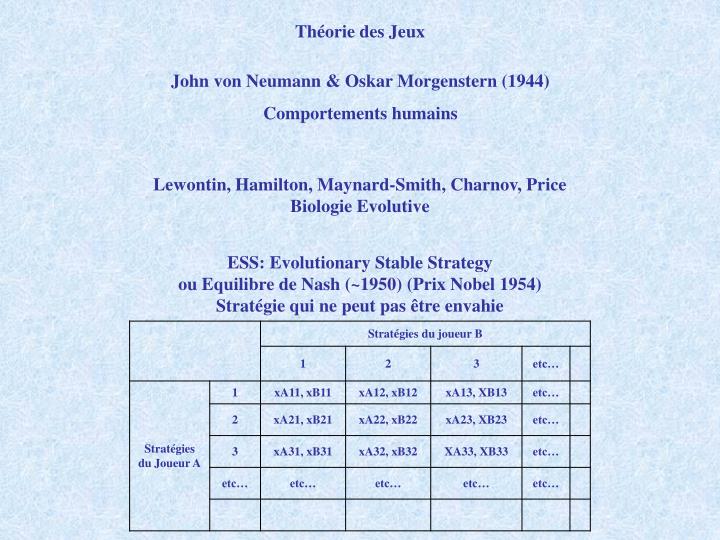
Théorie des Jeux. John von Neumann & Oskar Morgenstern (1944) Comportements humains. Lewontin, Hamilton, Maynard-Smith, Charnov, Price Biologie Evolutive. ESS: Evolutionary Stable Strategy ou Equilibre de Nash (~1950) (Prix Nobel 1954) Stratégie qui ne peut pas être envahie.

E N D
Théorie des Jeux John von Neumann & Oskar Morgenstern (1944) Comportements humains Lewontin, Hamilton, Maynard-Smith, Charnov, Price Biologie Evolutive ESS: Evolutionary Stable Strategy ou Equilibre de Nash (~1950) (Prix Nobel 1954) Stratégie qui ne peut pas être envahie
Dilemme du prisonnier Equilibre de Nash ESS Optimum de Pareto Vilfredo Pareto (1848-1923) sociologue Maximisation individuelle ≠ Maximisation du système Jeu sans équilibre
Jeux du faucon-colombe-bourgeois Maynar-Smith ESS si C<V Si C>V, une fréquence d'équilibre existe, f(faucon)=V/C Mixte envahit tout et n'est pas envahit→Mixte=ESS
Stratégie du bourgeois Si V>C faucon=ESS Si V<C bourgeois=ESS (mais n'envahit pas colombe)
Les bourgeois dans la vie Pararge aegeria (tircis) =>
V>C, ESS=Faucon V<C, Pas d'ESS 1) n=1 n<1+V/C, ESS=Faucon n≥1+V/C, ESS=Faucon et Tit For Tat Stratégie du Tit For Tat V>C V<C, ESS=Tit For Tat 2) n>1
Parasite Killer Diplomate Wh = Ph(Whmax-Rh) Wp = Pp(Wpmax-Vp) Wh = Whmax-Rh Wp = 0 Killer W Hôte WB Wh = 0 Wp = Wpmax-Vp Wh = Whmax-Gh Wp = Wpmax-Gp Diplomate Mutualisme (c<0) Commensalisme c=0 Wmax Parasitisme W = Wmax-c (c>0) Parasitoïdisme W = p(Wmax-c) p=f(c) c 0 Wmax
Vigilance des oiseaux (Maynard-Smith, 1989) Proportion de temps passé au nourrissage pour les stratèges x (mêmes stratèges cherchent en même temps) Probabilité d'être prédaté si non vigilant Gain par unité de temps de nourrissage k x*=ESS recherchée x le mutant qui ne peut envahir que si x=x* xx*=proportion de temps de non vigilance de tous les oiseaux kxx*=proportion de x prédatés x c W(x/x*)=Valeur sélective de x chez x*=cx(1-kxx*)=cx-ckx*x² On désire que W(x/x*) soit maximisé quand x=x*
Vigilance des oiseaux • W(x/x*)=Valeur sélective de x chez x*=cx(1-kxx*)=cx-ckx*x² • On désire que W(x/x*) soit maximisé quand x=x* • dW(x/x*)/dx=0 et d²W(x/x*)/dx<0 quand x=x* • dW(x/x*)/dx=c-2ckx*x=0 si x=x* • x*²=c/(2ck)(c≠0) • x*=1/√(2k) Si k=1/2, x*=1 Donc ici, si la probabilité de se faire prédater est de 1/2 l'ESS est de se nourrir 100% du temps (100% de non vigilance)
Dispersion des graines (Hamilton & May, 1977) n cases, une plante par case (1-m)N N graines produites par plante mN mNp p=survie à la dispersion
Dispersion des graines (Hamilton & May, 1977) n cases, une plante par case On cherche m* tel que si un mutant m survient dans une population m*, ce dernier ne peut envahir que si m=m* C'est-à-dire que W(m/m*) est maximum pour m=m* ou bien dW(m/m*)/dm=0 et d²W(m/m*)/dm<0 quand m=m* Case quelconque m* (n-1)Nm*p(1/n) + Nmp(1/n) + N(1-m*) graines soitenvirons, si n>>1 N(m*p+(1-m*)) graines en compétition Case de la plante mutante m (n-1)Nm*p(1/n) + Nmp(1/n) + N(1-m) soitenvirons, si n>>1 N(m*p+(1-m)) graines en compétition Dans la case m W(m/m*)=N (1-m)/( N(m*p+(1-m))) soitenvirons, si n>>1 W(m/m*) m=(1-m)/( m*p+(1-m) ) Dans les cases m W(m/m*)=(n-1)Nmp(1/n)/( N(m*p+(1-m*))) soitenvirons, si n>>1 W(m/m*)m*=mp/( m*p+(1-m*) ) W(m/m*)=mp/( m*p+1-m* ) + (1-m)/( m*p+1-m )
Dispersion des graines (Hamilton & May, 1977) W(m/m*)=mp/(m*p+1-m*) + (1-m)/( m*p+1-m) Rappel: (f/g)'=(f'g-fg')/g² On veut que dW(m/m*)/dm=0 pour m=m*, soit Si p≠0 et m*≠1/(1-p) et donc p≠1 m*p+1-m*-m*=0 m*(p-2)+1=0 m*=1/(2-p) Donc même si la survie à la dispersion p est proche de 0, l'ESS m*≈1/2






















