Download

1 / 52
650 likes | 2.04k Views
เอนไซม์ (Enzyme). บริษัท ไทย เซมเทค จำกัด. To protect the environment. เทคโนโลยีชีวภาพ เพื่อ รักษาสิ่งแวดล้อม. www.zymetec.com. 02-450 6038. Made in USA. ความเป็นมาของ เอนไซม์.

E N D
เอนไซม์ (Enzyme) บริษัท ไทยเซมเทค จำกัด To protect the environment.เทคโนโลยีชีวภาพเพื่อรักษาสิ่งแวดล้อม www.zymetec.com 02-450 6038 Made in USA.
ความเป็นมาของ เอนไซม์ เอนไซม์ถูกนำมาใช้ครั้งแรก ในปี ค.ศ. 1878 ซึ่งมีการใช้ในการเร่งปฏิกิริยาการหมักน้ำตาลของยีสต์ พบว่าสามารถเร่งปฏิกิริยาให้เร็วขึ้น คำว่า Enzyme มาจากภาษากรีก แปลว่า In yeast - เอนไซม์เป็นโปรตีนที่มีลักษณะก้อนกลม ที่ถูกสังเคราะห์ขึ้นเพื่อทำหน้าที่ตัวเร่งปฏิกิริยาเคมีต่างๆในกระบวนการเมตาบอลิซึมของสิ่งมีชีวิต โดยมีการทำงานที่คล้ายคลึงกันกับการทำงานของคะตะลิสต์
เอนไซม์ (Enzyme) เอนไซม์ คือ สารที่เร่งปฏิกิริยาโดยทำหน้าที่เป็น catalyst มีความจำเพาะกับ substrate สูง โดยเอนไซม์ทำงานเร่งปฏิกิริยาโดยลดพลังงานกระตุ้น ส่วนใหญ่เป็นสารอินทรีย์กลุ่มโปรตีน แบ่งเป็น 2 ชนิด คือ 1. เอนไซม์ที่ทำงานภายในเซลล์ (endoenzyme หรือ intracellular enzyme) 2. เอนไซม์ที่ทำงานภายนอกเซลล์ (exoenzyme หรือ extracellular enzyme) เอนไซม์ประกอบด้วยส่วนที่เป็น โปรตีน หรือ apoenzyme รวมกับสารอินทรีย์ ที่เรียกว่า coenzyme จึงเป็นเอนไซม์ที่สมบูรณ์ หรือ holoenzyme ในบางครั้งอาจมีไอออนที่สำคัญในการทำงานของเอนไซม์เรียกว่า cofactor การทำงานของเอนไซม์
ลักษณะพิเศษของเอนไซม์ที่แตกต่างจากคะตะลิสต์ลักษณะพิเศษของเอนไซม์ที่แตกต่างจากคะตะลิสต์ • มีความจำเพาะ (selectivity) ต่อปฏิกิริยาและซับสเตรท โดยที่เอนไซม์หนึ่งจะทำงานได้ขึ้นอยู่กับโครงสร้างของซับสเตรทที่จะสามารถจับได้พอเหมาะภายในร่อง • มีความสามารถในการเร่งได้สูงมาก โดยบอกเป็นค่าหมุนเวียน (turnover number) ซึ่งหมายถึง จำนวนสารตั้งต้นที่ทำปฏิกิริยาเปลี่ยนเป็นผลิตผลของปฏิกิริยาใน 1 หน่วยเวลา เมื่อใช้เอนไซม์ 1 โมเลกุล โดยเอนไซม์ต่างๆจะมีค่านี้อยู่ระหว่าง 10 – 108 ต่อวินาที
โครงสร้างของเอนไซม์ เอนไซม์มีขนาดตั้งแต่ 12,000 ถึง กว่า 1 ล้านดาลตัน ส่วนใหญ่เป็นสารประกอบโปรตีน โครงสร้างของเอนไซม์แบ่งได้ 2 ชนิด คือ • โปรตีนธรรมดา (simple protein) เป็นโปรตีนที่มีกรดอะมิโนเป็นองค์ประกอบอย่างเดียว เอนไซม์พวกนี้มีโครงสร้างเป็นทรงกลม • โปรตีนรวมตัวกับสารอื่น (conjugated protein) ประกอบด้วยกรดอะมิโนรวมกับสารอื่นที่ไม่ใช่โปรตีน ที่เรียกว่าโคแฟกเตอร์ (cofactor) จึงจะสามารถทำงานได้อย่างสมบูรณ์ เรียกเอนไซม์กลุ่มนี้ว่า holoenzyme ถ้าแยกโคแฟกเตอร์ออกจะทำให้เอนไซม์กลุ่มนี้จะไม่สามารถทำงานได้อย่างสมบูรณ์เรียกว่า อะโพเอนไซม์ (apoenzyme)
Apoenzyme + Cofactor = Holoenzyme Inorganic เช่น Fe 2+ ion etc. Organic เช่น vitamin เรียก coenzyme
โคแฟกเตอร์ แบ่งได้เป็น 2 พวกคือ • สารอนินทรีย์หรือไอออนของโลหะ (metal ions) หรือเกลือแร่มีโครงสร้างง่ายๆได้แก่ Fe 2+ , Mg 2+ , Mn 2+เป็นต้น • สารอินทรีย์หรือโคเอนไซม์ เป็นโมเลกุลอินทรีย์มีโครงสร้างซับซ้อน ที่มีขนาดเล็กและทนต่อความร้อนได้ดี โคเอนไซม์ส่วนใหญ่จะเป็นวิตามินที่ละลายน้ำ ทำหน้าที่ในการย้ายหมู่ต่างๆ เช่น หมู่ไฮโดรเจน อะมิโน ฟอสเฟต โดยถ้า โคเอนไซม์จับกับอะโพเอนไซม์อย่างหลวมๆ เรียก โคเอนไซม์กลุ่มนี้ว่า ซับสเตรทร่วม (cosubstrate) แต่ถ้าโคเอนไซม์จับกันอย่างแน่นหนากับเอนไซม์ด้วยพันธะโควาเลนต์ จะถูกเรียกว่าหมู่พรอสเทติก (prostetic group)
เอนไซม์ที่ผลิตโดยร่างกาย แบ่งได้เป็น 2 กลุ่ม คือ • เอนไซม์ย่อยอาหาร (digestive enzyme) ทำหน้าที่ย่อยอาหารภายในระบบทางเดินอาหารเพื่อส่งสารอาหารไปยังเซลล์ต่างๆในร่างการ ได้แก่ 1.1 เอนไซม์โปรตีเอส (protease) ย่อยโปรตีน 1.2 เอนไซม์ไลเปส (Lipase) ย่อยไขมัน 1.3 เอนไซม์อะไมเลส (amylase) ย่อยแป้ง 2. เมตาบอลิคเอนไซม์ (metabolic enzyme) มีอยู่ในเซลล์ทุกเซลล์ในทุกอวัยวะ เอนไซม์นี้จะทำหน้าที่เกี่ยวกับทุกปฏิกิริยาที่เกิดขึ้นในร่างกาย มีหน้าที่ซ่อมแซมส่วนที่สึกหรอ ช่วยระบบภูมิคุ้มกัน
เอนไซม์แบ่งได้ 2 ประเภทคือ • ไอโซเอนไซม์ หรือ ไอโซไซม์ (isoenzyme หรือ isozyme) เป็นเอนไซม์ที่มีโครงสร้างแตกต่างกันแต่สามารถเร่งปฏิกิริยาเดียวกันได้ • อัลโลสเตอริกเอนไซม์ (allosteric enzyme) หรือ เอนไซม์ควบคุม (regulatory enzyme) เป็นเอนไซม์ที่ประกอบด้วยหน่วยย่อยๆหลายหน่วยร่วมกัน เช่น หน่วยย่อยเร่ง (catalytic enzyme) สารที่มีอิทธิพลต่อการควบคุมการทำงานของเอนไซม์เหล่านี้ เรียกว่า ตัวควบคุม (effector หรือ modulator หรือ regulator) ตัวควบคุม มี 2 ชนิด คือ 2.1 ตัวเร่ง (activator) ทำให้เอนไซม์ทำงานได้ดีขึ้น เรียก positive modulator 2.2 ตัวยับยั้ง (inhibitor) ทำให้เอนไซม์ทำงานได้ช้าลง เรียกว่า negative modulator
บริเวณของเอนไซม์ที่มีตัวควบคุม เรียก บริเวณควบคุม (regulatory site หรือ allosteric site) • เรียก เอนไซม์ที่มีทั้งบริเวณเร่งและบริเวณควบคุมว่า อัลโลสเตอริกเอนไซม์
การเรียกชื่อตามแบบสามัญ (recommended name) มีหลักเกณฑ์ดังนี้คือ • เรียกตามชื่อของซับสเตรทที่เอนไซม์เร่งปฏิกิริยา แล้วลงท้ายด้วยคำว่า “ase”เช่น เอนไซม์เร่งปฏิกิริยาการสลายยูเรีย เรียกว่าเอนไซม์ยูรีเอส (Urease) • เรียกตามชนิดของปฏิกิริยาแล้วลงท้ายด้วยคำว่า “ase”เช่นเอนไซม์ออกซิเดส (oxidase) เร่งปฏิกิริยาออกซิเดชันของซับสเตรทที่มี O2 • เรียกตามข้อที่ 1 และ 2 รวมกัน เช่น ไกลซีนดีคาร์บอกซิเลส (glycinedecarboxylase) • เรียกตามชื่อเฉพาะไม่สัมพันธ์กับซับสเตรทหรือปฏิกิริยาที่เกี่ยวข้อง เช่น เพปซิน (pepsin) เป็นเอนไซม์ย่อยโปรตีน เป็นต้น
การเรียกชื่อเอนไซม์ตามระบบ (systemic name) ตามข้อตกลงของกรรมาธิการเอนไซม์นานาชาติ ได้มีการเรียกชื่อตามตัวเลข โดยมีลักษณะคือ EC xx.xx.xx.xx (อักษร x คือตัวเลข) เช่น EC 2.7.3.2 2 = ชนิดเอนไซม์ transferase 7 = ชนิดย่อยของรหัสตัวที่ 1 หมายถึงการย้ายหมู่ฟอสเฟต 3 = ชนิดย่อยของรหัสตัวที่ 2 หมายถึงการแบ่งย่อยของหมู่ฟอสเฟต 2 = รหัสเฉพาะตัวของเอนไซม์ชนิดนั้น Enzyme functional class Sub sub class ลำดับที่ subclass
การแบ่งเอนไซม์ตามชนิดของปฏิกิริยาการแบ่งเอนไซม์ตามชนิดของปฏิกิริยา • oxidoreductase ปฏิกิริยาออกซิเดชัน-รีดักชัน การถ่ายทอด • อิเล็กตรอน • 2. transferase ย้ายหมู่ฟังก์ชันจากสารหนึ่ง ไปอีกสารหนึ่ง • 3. hydrolase ไฮโดรไลซิส แยกพันธะด้วยน้ำ • 4. lyases เติมพันธะคู่ • 5. isomerase ไอโซเมอร์ไรเซชัน • 6. ligases สร้างพันธะโดยสลาย ATP รูปร่างโมเลกุลของเอนไซม์
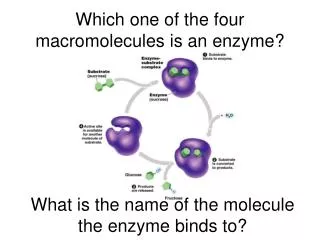
กลไกการทำงานของเอนไซม์กลไกการทำงานของเอนไซม์ ให้ E = Enzyme S = Substrate P = Product ES = Enzyme เมื่อจับกับ Substrate EP = Enzyme เมื่อผลิต Product และยังคงจับกันอยู่
การทำงานของเอนไซม์ ขั้นตอนแรก : เอนไซม์ E รวมตัวกับสารตั้งต้น S ได้เป็นสารประกอบ ES E + S ES ขั้นตอนที่สอง : สารประกอบ ES เกิดการเปลี่ยนแปลงเป็น ES* ES* EP
การทำงานของเอนไซม์ ขั้นตอนที่สาม :เกิด สารประกอบ ระหว่างเอนไซม์กับผลผลิต ขั้นตอนที่สุดท้าย :ได้ผลผลิตออกมา และได้เอนไซม์กลับมาเหมือนเดิม EP E + P
จลน์ศาสตร์ของเอนไซม์ E + S ES E + P k1 k2 k 3 k 4
จลนศาสตร์ของเอนไซม์ • เอนไซม์ทำปฏิกิริยากับสับสเตรท เกิดเป็นเอนไซม์-สับสเตรทคอมเพลกซ์ก่อนแล้วจึงสลายในขั้นตอนที่ 2 ให้ผลผลิต และเอนไซม์กลับคืนมา อัตราเร็วของปฏิกิริยาในการเปลี่ยน S ไปเป็น P ขึ้นอยู่กับอัตราเร็วในการ เปลี่ยน ES ไปเป็น P และ E นั่นคือ อัตราเร็วในการเกิด P จะขึ้นอยู่กับความเข้มข้นของ ES • เมื่อ k1 k2 k3 และ k4 คือ ค่าความเร็วคงที่ (velocity constant) หรือ ค่าอัตราเร็วคงที่ (rate constant) ของปฏิกิริยาทั้ง 4 • ที่ steady state อัตราเร็วในการเกิด ES จะเท่ากับอัตราเร็วในการสลาย ES ทำให้ [ES] คงที่ จะได้
สมการไมคีลิส-เมนเทน (Michaelis-Menten constant) • แสดงถึง ความสัมพันธ์ระหว่างอัตราเร็วของปฏิกิริยา และความเข้มข้นของ S เมื่อทราบ Vmax และ KM • Vmaxคือ ความเร็วสูงสุด (maximum velocity) ของการทำงานของเอนไซม์ • KM คือ ค่าคงที่ไมคีลิส-เมนเทน (Michaelis-Menten constant) ของปฏิกิริยาของเอนไซม์ที่ steady state มีหน่วยเป็นโมล/ลิตร • V = Vmax[S] • KM + [S] • ที่ V = Vmax /2 หรือ ครึ่งหนึ่งของความเร็วสูงสุด (half maximal) จะได้ KM = [S]
สมการไลน์วีเวอร์-เบอร์กสมการไลน์วีเวอร์-เบอร์ก • เป็นสมการจากการกลับสมการไมคีลิส-เมนเทน • 1 = KM . 1 + 1 • v Vmax [S] Vmax • เมื่อเขียนกราฟระหว่าง 1/v และ [S] จะได้กราฟเส้นตรงที่มีความชัน = KM/Vmax และจุดตัดแกน 1/v คือ 1/Vmax และจุตัดแกน 1/[S] คือ -1/KM ทำให้สามารถหาค่า KM และ Vmax ที่ถูกต้องได้
Enzyme Kinetics Student A Enzyme activity Score Student B Student C 0 1 2 3 4 0 1 2 3 4 Exam Chapters Substrate concentration Increase the substrate concentration, observe the change of enzyme activity Juang RH (2004) BCbasics
Increase Substrate Concentration 0 1 2 3 4 5 6 7 8 80 60 40 20 0 Product 0 2 4 6 8 Substrate (mmole) S + E ↓ P (in a fixed period of time) Juang RH (2004) BCbasics
Essential of Enzyme Kinetics E S Steady State Theory E E + + P S In steady state, the production and consumption of the transition state proceed at the same rate. So the concentration of transition state keeps a constant. Juang RH (2004) BCbasics
Constant ES Concentration at Steady State Concentration Reaction Time S P ES E Juang RH (2004) BCbasics
An Example for Enzyme Kinetics (Invertase) 1 vo vo 1 Vmax - 1 Km 1/S S 1)Use predefined amount ofEnzyme→E 2)Add substratein various concentrations →S(x ,y) 3)MeasureProductin fixedTime (P/t) →vo(y, y) 4)(x, y)plot get hyperbolic curve,estimate→Vmax 5)Wheny = 1/2 Vmaxcalculatex ([S]) →Km Vmax 1/2 Juang RH (2004) BCbasics Km Double reciprocal Direct plot
A Real Example for Enzyme Kinetics Substrate Product Velocity Double reciprocal 2.0 1.0 0 1.0 0.5 0 v 1/v 1.0 -3.8 -4 -2 0 2 4 0 1 2 [S] 1/[S] Data v (mmole/min) [S] Absorbance 1/S 1/v no 0.25 0.50 1.0 2.0 0.21 0.36 0.40 0.46 0.42 0.72 0.80 0.92 4 2 1 0.5 2.08 1.56 1.35 1.16 1 2 3 4 → → → → (1) The product was measured by spectroscopy at 600 nm for 0.05 per mmole (2) Reaction time was 10 min Direct plot Double reciprocal Juang RH (2004) BCbasics
การยับยั้งแบบถาวร ปัจจัยที่ควบคุมการทำงานของเอนไซม์ 1.ความเข้มข้นของเอนไซม์ 2.ความเข้มข้นของสารตั้งต้น หรือซับสเตรต (substrate) 3.ค่าความเป็นกรด-เบส (pH) 4.อุณหภูมิ (optimum temperature) การยับยั้งการทำงานของเอนไซม์ - การยับยั้งแบบถาวร Inhibiter ทำให้ส่วน active site เปลี่ยนแปลงไป เอนไซม์ไม่สามารถกลับสู่ในสภาพที่ทำงานได้อีก - การยับยั้งที่กลับคืนได้ 1.การยับยั้งแบบแข่งขัน โดยตัวยับยั้งมีโครงสร้างคล้ายกับซับสเตรตทำให้ สามารถแย่งจับกับเอนไซม์ได้ 2.การยับยั้งแบบไม่แข่งขัน โดยสารเคมีบางชนิดไปจับกับส่วน cofactor ทำให้เอนไซม์ไม่สามารถทำงานได้ การยับยั้งแบบไม่แข่งขัน การยับยั้งแบบแข่งขัน
ปัจจัยที่มีผลต่อการทำงานของเอนไซม์ปัจจัยที่มีผลต่อการทำงานของเอนไซม์ • 1. อุณหภูมิ เมื่ออุณหภูมิสูงขึ้น แอกติวีตีของเอนไซม์จะเพิ่มขึ้นเรื่อยๆ จนถึงจุดสูงสุดหนึ่ง (temperature optimum) แล้วจะลดลง ซึ่งจำเพาะตามชนิดของเอนไซม์ ส่วนใหญ่อยู่ในช่วงอุณหภูมิ 25–40oC แต่ที่อุณหภูมิ สูงกว่าจุดนี้มากๆ เอนไซม์จะเกิดการเสียแอกติวีตีทางชีวภาพ • 2. pH จุดที่ทำให้เอนไซม์มีแอกติวีตีสูงสุด เรียกว่า pH optimum ซึ่งส่วนใหญ่อยู่ในช่วง pH 6-7.5 ถ้า pH สูง หรือต่ำเกินไปจะทำให้เกิดการสูญเสียแอกติวีตีทางชีวภาพ • 3. ความเข้มข้นของเอนไซม์ ที่ความเข้มข้นของสับสเตรทมากเกินพอ (excess) อัตราเร็วของปฏิกิริยา จะเพิ่มขึ้น เมื่อความเข้มข้นของเอนไซม์เพิ่มขึ้น • 4. ความเข้มข้นของสับสเตรท เมื่อความเข้มข้นของเอนไซม์คงที่ ที่ความเข้มข้นของสับสเตรทน้อยๆ ปฏิกิริยาจะ เกิดขึ้นด้วยอัตราเร็วมากจนกระทั่งถึงความเข้มข้นของสับสเตรทจุดหนึ่งที่อัตราเร็วของ ปฏิกิริยาจะคงที่ ความเร็วของปฏิกิริยาที่สูงสุด เรียกว่า ความเร็วสูงสุด (Vmax)
ตัวยับยั้งเอนไซม์ แบ่งออกได้เป็น 2 ประเภท 1. ตัวยับยั้งแบบผันกลับไม่ได้ (irreversible inhibitor) เกี่ยวกับการทำลายหรือเปลี่ยนแปลง functional group 1 หมู่หรือมากกว่าที่จำเป็นสำหรับแอกติวีตีบนโมเลกุลของเอนไซม์ ซึ่งตัวยับยั้งแบบนี้จะจับ กับเอนไซม์อย่างแน่นทั้งแบบโควาเลนท์และนอนโควาเลนท์ เช่น การยับยั้งเอนไซม์ไซโคลออกซิจิเนส (cyclooxygenase) โดยยาแอสไพริน (aspirin หรือ acetyl salicylate) 2. ตัวยับยั้ง แบบผันกลับได้ (reversible inhibitor) แบ่งเป็น 3 ชนิด
2.1 ตัวยับยั้งแบบแข่งขัน (competitive inhibitor) หมายถึง สารที่สามารถเข้า ไปแย่งสับสเตรทจับที่บริเวณแอกตีฟบนโมเลกุลของเอนไซม์ แล้วทำให้แอกติวีตีในการเร่งปฎิกิริยา ของเอนไซม์ลดลง โดยค่า KM เปลี่ยน แต่ Vmax คงที่ เช่น กรดมาโลนิกสามารถจับเอนไซม์ซักซินิก ดีไฮโดรเจเนส (succinic dehydrogenase) ในปฏิกิริยาการออกซิไดส์กรดซักซินิกไปเป็นกรด ฟูมาริก เพราะมีโครงสร้างคล้ายกับกรดซักซินิก
2.2 ตัวยังยั้งแบบไม่แข่งขัน (noncompetitive inhibitor) หรือตัวยับยั้ง แบบผสม (mixed inhibitor) โดยตัวยับยั้งชนิดนี้จะเข้าไปจับที่บริเวณอื่นบนโมเลกุลของ เอนไซม์ที่ไม่ใช่บริเวณเร่ง เช่นบริเวณควบคุม ทำให้โครงร่างของเอนไซม์เปลี่ยนแปลงไป ทำให้บริเวณเร่งเปลี่ยนไปจึงทำงานไม่ได้ เช่น Cu2+Hg2+ และ Ag+ หรืออนุพันธ์ (derivative) ของมันสามารถทำปฏิกิริยากับ หมู่-SH ของซิสเตอีนทำให้โครงรูปสามมิติ (three dimensional conformation) ที่จำเพาะของเอนไซม์เสียไป ในกรณี noncompetitive inhibitor ค่า Vmax เปลี่ยนแปลง(ลดลง) ส่วน KM ไม่เปลี่ยนแปลง
2.3 การยับยั้งแบบอันคอมเพติตีฟ (uncompetitive inhibition) การยับยั้ง แบบนี้เกิดขึ้นเมื่อตัวยับยั้งเข้ารวมเฉพาะกับเอนไซม์สับสเตรทคอมเพลกซ์เท่านั้นแบบไม่ผันกลับ เกิดเป็น ESI คอมเพลกซ์ ซึ่งจะไม่ได้ผลผลิตเกิดขึ้น ลักษณะเหมือนกับการยับยั้งแบบไม่แข่งขัน โดยไม่เกิดการ ผันกลับ (reversible) เมื่อเพิ่มความเข้มข้นของสับสเตรท ค่า Vmax ลดลง และค่า Km ลดลง แต่ความชันไม่เปลี่ยนแปลง
Enzyme Inhibition (Mechanism) E + S→ES→E + P + I ↓ EI E + S→ES→E + P + + II ↓ ↓ EI+S→EIS E + S→ES→E + P + I ↓ EIS ← ← ← ↑ ↑ ↑ ↑ Uncompetitive Competitive Non-competitive E Substrate E X Cartoon Guide Compete for active site Inhibitor Different site Equation and Description [I] binds to free [E] only, and competes with [S]; increasing [S] overcomes Inhibition by [I]. [I] binds to [ES] complex only, increasing [S] favors the inhibition by [I]. [I] binds to free [E] or [ES] complex; Increasing [S] can not overcome [I] inhibition. Juang RH (2004) BCbasics
Enzyme Inhibition (Plots) Uncompetitive Competitive Non-competitive Vmax Vmax vo Vmax’ Vmax’ I Direct Plots Km [S], mM Km’ Km [S], mM 1/vo 1/vo 1/vo I I Double Reciprocal Two parallel lines Intersect at X axis Intersect at Y axis 1/Vmax 1/Vmax 1/Vmax 1/Km 1/[S] 1/Km 1/[S] 1/Km 1/[S] Vmax vo I I Km Km’ [S], mM =Km’ Vmax unchanged Km increased Vmax decreased Km unchanged Both Vmax & Km decreased I Juang RH (2004) BCbasics
หน่วยวัดการทำงานของเอนไซม์หน่วยวัดการทำงานของเอนไซม์ • หน่วยเอนไซม์ หมายถึง จำนวนเอนไซม์ที่ทำให้ปริมาณของผลิตภัณฑ์เกิดขึ้นหรือปริมาณของซับสเตรทลดลง มีค่าเป็น ไมโครโมลต่อนาที หน่วยสากล (international unit, IU หรือ U) 1.0 IU คือ แอคติวิตีของเอนไซม์หรือปริมาณของเอนไซม์ที่เร่งการเปลี่ยน 1.0 ไมโครโมลของซับสเตรทใน 1 นาทีที่อุณหภูมิ 25 º C คาทาล (katal : kat) หมายถึง ปริมาณของเอนไซม์ที่เร่งการเปลี่ยนแปลง 1 โมลของซับสเตรทใน 1 วินาที หน่วยที่นิยมใช้ คือ µkat , nkat, pkat 1 kat = 1/60 U = 1.67 x 10 -2 U 2. หน่วยแอคติวิตีเฉพาะ (specific activity) หมายถึง หน่วยเอนไซม์ ต่อ 1 มิลลิกรัมของโปรตีน ใช้ในการหาความบริสุทธิ์ของเอนไซม์
Enzyme Kinetics Direct plot Significance kcat/Km zero order 1st order Vmax[S] kcat vo= E3 Km+[S] Turn over number E2 E1 k3 [Et] Competitive Double reciprocal Activity Unit 1 mmole min Non-competitive Specific Activity unit mg Uncompetitive Observe vo change under various [S], resulted plots yield Vmax and Km Vmax Km & Maximum velocity Affinity with substrate Inhibition Bi-substrate reaction also follows M-M equation, but one of the substrate should be saturated when estimate the other Activity Juang RH (2004) BCbasics
Significance of Enzyme Kinetics Vmax[S] vo = Km+[S] v0 = Vmax × K = k3 [Et]× K ObtainVmaxandKm zero order E3 Proportional to enzyme concentration E2 E1 1st order [S] = Low → High [S] = Fixed concentration Juang RH (2004) BCbasics
Km: Affinity with Substrate Vmax[S] vo = Km+[S] Vmax Ifvo = 2 Vmax Vmax[S] = 2 Km+[S] Km+[S] = 2[S] Km= [S] Km When using different substrate Vmax S2 S1 S3 1/2 S1S2S3 Juang RH (2004) BCbasics Affinity changes
Km: Hexokinase Example CHO H-C-OH HO-C-H H-C-OH H-C-OH H2-C-OH CHO H-C-OH H-C-OH H-C-OH H-C-OH H2-C-OH CHO HO-C-H HO-C-H H-C-OH H-C-OH H2-C-OH Glucose + ATP → Glc-6-P + ADP Substrate Glucose Allose Mannose number 1 2 3 4 5 6 Km = 8 8,000 5 mM Juang RH (2004) BCbasics
Turn Over Number, kcat k1 k3 E + S ES E + P (vo) k2 Second order (IV) k3 Vmax[S] k3 [E][S] vo = = = [E][S] Km+[S] Km+[S] Km Omit the [substrate] Substrate specificity When substrate excess,k3 =kcat, turn over number (t.o.n) When [substrate] is low Start from M-M eq. Juang RH (2004) BCbasics