Download

1 / 46
470 likes | 700 Views
第七章 细胞因子 CYTOKINE ). 概念. 由机体免疫系统的细胞产生、分泌,通过细胞间的信号传导调节免疫应答的蛋白质。. 特性. 一种细胞因子可由多种细胞产生,单一刺激也可使同一种细胞产生多种细胞因子。 均为分子量小于 8000 的小分子量分泌型糖蛋白。 几乎都是细胞在受抗原或丝裂原刺激后的活化过程中合成和分泌,并通过自分泌( autocrine )、旁分泌( paracrine )和内分泌( endocrine )的方式发挥作用。. 细胞因子的命名. 白细胞介素( interleukins , IL)

E N D
第七章 细胞因子CYTOKINE) 概念 由机体免疫系统的细胞产生、分泌,通过细胞间的信号传导调节免疫应答的蛋白质。
特性 • 一种细胞因子可由多种细胞产生,单一刺激也可使同一种细胞产生多种细胞因子。 • 均为分子量小于8000的小分子量分泌型糖蛋白。 • 几乎都是细胞在受抗原或丝裂原刺激后的活化过程中合成和分泌,并通过自分泌(autocrine)、旁分泌(paracrine)和内分泌(endocrine)的方式发挥作用。
细胞因子的命名 • 白细胞介素(interleukins,IL) • 白细胞介素是调节淋巴细胞与其他白细胞之间作用的细胞因子,其界定范围很广,除共用白细胞介素这一相同名称外,实际上是不同性质蛋白质的混合群体,根据发现的先后而命名,现已报道有23种IL,
2. 干扰素(interferons,IFN) 由病毒感染、免疫及其它各种化学诱导剂诱导机体产生的具有抑制病毒感染、免疫刺激及其他多种化学刺激作用的糖蛋白。往往通过干扰病毒RNA和蛋白的合成而抑制病毒感染,已发现有5中不同类型的IFN。
3. 肿瘤坏死因子 (Tumor necrosis factors,TNF) 具有能使肿瘤细胞死亡作用的细胞因子,主要包括两种。
4. 生长因子(Growth factors) 能刺激细胞生长的细胞因子,在保证机体自身细胞的更新过程中起重要作用。
5. 趋化因子(Chemokines) 一群主要在炎症过程中发挥作用的小分子量蛋白质。如IL-8
细胞因子的总体功能 1. 细胞因子的作用途径 细胞因子可在多个靶细胞上发挥作用,主要通过自分泌(autocrine)、旁分泌(paracrine)和内分泌(endocrine)三种途径发挥作用。
2. 细胞因子的功能特点 细胞因子作用于靶细胞后,能导致细胞分裂、分化,或刺激细胞表达新的受体或其他蛋白质产物,相反也可抑制上述功能。 a. 多效性(pleiotropy) 大多数细胞因子可作用于多种靶细胞,并在不同的靶细胞产生不同的效应。 b. 过量性(redundancy) 多种细胞因子作用于同一种靶细胞,且往往量大。如B细胞的功能受IL-2、IL-5、IL-4和IL-6等细胞因子的共同作用。
c. 协同性(synergy) 有的细胞因子的作用与其他的细胞因子之间有密切的相关性。多发生在两种或两种以上细胞因子同时共同作用于一个靶细胞时,如B细胞的功能受IL-5和IL-4的共同作用开启IgE产生的开关;也可以以顺序的方式发挥作用,即一种细胞因子刺激靶细胞产生另一种细胞因子的受体,从而增强另一种细胞因子的作用, d. 抵消性(antagonizing) 即一种细胞因子的作用可被另一种所抵消。如IFN-γ 能抑制IL-4刺激B细胞产生IgE的能力。
细胞因子的结构 根据对细胞因子结构的分析,可将细胞因子分为四大群。 第一群 IL-2、3、4、5、6、7、9、10、11、13、15 、21 、30 、G-CSF、M-CSF、GM-CSF和IFN等细胞因子属于该群,该群的蛋白质分子由四个α螺旋聚合在一起,根据链的长短可分为两个亚群。IL-2、3、4、7、9属短链,IL-6、10、11属长链。具有免疫调节和造血功能。
第二群 IL-1、TNF、TGF- β等细胞因子属于该群,该群的蛋白质分子有一长链的β 折叠。其功能主要为生长作用、调节组织细胞的衰老和死亡,并与机体的应激有关。 第三群 IL-8等细胞因子属于该群,该群的蛋白质分子具α螺旋和β 折叠的小分子蛋白质。与炎症反应有关。
第四群 IL-12等细胞因子属于该群,该群的蛋白质分子具有不同结构单位的镶嵌混合物。其功能取决于装配该分子的基本成分,如IL-12是第一群结构的嵌合体,功能与第一群细胞因子相同。
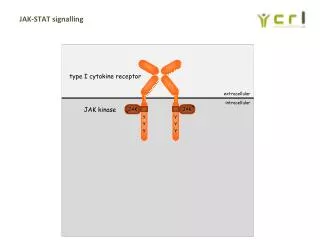
细胞因子的受体 细胞因子的受体是整合在细胞膜上的膜蛋白,往往由膜内区和膜外区两个功能区组成,膜外区与细胞因子结合,而膜内区则参与信号传导。 根据其结构和活性可分为四个家族。 第一家族 细胞因子/干扰素受体家族,是最大的受体家族,包括IL-2Rβ、IL-3R、IL-4R、IL-5R α 、IL-7Rα、IL-9R、IL-12R、 G-CSFR、GM-CSFR、IFNα/βR、IFN-γR和IL-10R,还包括共同β链的IL-3、IL-5受体和共同γ链的IL-2、IL-4、IL-9、IL-15的受体。在结构上有一个或两个包含特定性的膜外区,具有完整的胱氨酸残基。
第二家族 受体激酶家族,TGF-βR属此类受体,在结构上具有共同的膜内区。通常情况下以单体存在,活化后成为双体结构。 第三家族 TNF/神经生长受体家族,TNF- αR和TNF-βR属此类受体,在结构上具有一个富含胱氨酸的单折叠跨膜区,而膜内区则极少有同原性,因此,这种受体的配体的生物学活性差异很大,能启动细胞凋亡的fas(CD95)和CD40L也能与之结合,促使细胞凋亡。
第四家族 七折受体家族,是包括IL-8在内的趋化因子的受体,同时也是C5a和血小板活化因子(PAF)D的受体。其结构特征是在细胞膜上有七个折。其信号传导需G蛋白介导。
细胞因子的调节 细胞因子可通过受体表达、受体排斥、特异蛋白结合及相反功能细胞因子的抑制等方式进行调节其功能。如T细胞对IL-2的反应主要取决于靶细胞表面IL-2R的表达;IL-1的作用主要受受体抵消物(IL-1RA)的调节;另一些细胞因子则被体液中的特异性可溶性蛋白质所结合而失去其应有的功能,这些可溶性蛋白主要是游离在体液中的细胞因子受体,如IL-2R、TNFR-I、TNFR-II、IL-6R等受体已证明可成为可溶性的物质而游离于体液中;相反功能的细胞因子的相互作用是调节细胞因子作用的主要方式。
Interleukin-1 除巨噬细胞外,Langerhans 、T、B、NK细胞、血管内皮细胞、纤维母细胞及角质细胞等也能产生,IL-1有三种不同的产物:IL-1α、IL-1β和IL-1RA。 IL-1主要作用于T、B、NK、中性粒细胞、碱性粒细胞、树突状细胞、纤维母细胞、内皮细胞、角质细胞和单核细胞, IL-1α、IL-1β是Th2细胞的共刺激因子,也是主要的应激反应刺激因子。 IL-1RA是没有生物学活性的蛋白,能与IL-1的受体结合,但不能产生任何刺激信号,因此是IL-1的拮抗物。
Interleukin-6 除巨噬细胞外,T、B细胞、骨髓基质细胞(marrow stromal cells)、血管内皮细胞、纤维母细胞及角质细胞等也能产生。 IL-6主要作用于T、B、骨髓基质细胞。 能促进IL-2和IL-2R的产生及T细胞的分化,在促进IgM和IgA合成过程中是IL-1和IL-5的协同因子。此外,还能诱导肝细胞产生急性反应蛋白(acutephase proteins),并具有热原(pyrogen)的作用。
Interleukin-12 主要由活化的巨噬细胞产生,通过诱导IL-2和IFN-γ的分泌而促进Th1细胞的活性及增强T细胞和NK细胞的分化与细胞毒性,并能通过IL-4的合成抑制IgE的产生,可增强B细胞分泌免疫球蛋白的能力,与TNF-α一起诱导LAK细胞(lymphokine-activated killer cells)的产生。
TNF-α 除巨噬细胞外,T细胞、B细胞和成纤维细胞也能产生,几乎能作用于所有的真核细胞,是许多免疫反应和炎症反应的介质,能调节许多种类细胞的生长,并具有触发某些肿瘤细胞的溶解。
主要由T细胞产生的细胞因子 1. 主要由Th1细胞产生的细胞因子
Interleukin-2 主要由T细胞及部分B细胞产生,具有广谱的免疫增强作用,在免疫应答中起关键作用,可诱导T、B细胞的增殖分化,促进细胞毒性T细胞前体细胞分化为细胞毒性T细胞,增强其杀伤效应,并可促进NK细胞的功能与释放免疫干扰素,此外,还具有显著的抗肿瘤和抗病毒作用。 IL-2受体是一个由α、β和γ三条链组成的异聚体, α链与该受体的亲和力密切相关。
IFN-γ 除Th1细胞外,CD8+T细胞和NK细胞也能产生,作用于T、B、NK和巨噬细胞。刺激B细胞产生IgG2a,对B细胞产生IgG3、IgG1、IgG2b的作用较弱,增强T细胞表达MHC I类分子,诱导Th1细胞产生IL-2及其受体,能作用于Th2细胞抑制IL-4的产生,从而阻断IgE的产生。增强NK细胞的功能,激活巨噬细胞,极大地增强其消化和处理被吞噬的微生物的能力,促进抗体介导的吞噬作用和ADCC作用。 能增加肿瘤细胞、病毒感染细胞及抗原提呈细胞等的MHC I和MHC II类分子的表达。提高单核细胞表达MHC III类产物的表达。
Tumor Necrosis Factor-β,TNF-β 主要有Th1细胞和活化的CD8+T细胞产生。可以被分泌成可溶性状态,也可能与淋巴毒素(lymphotoxin-β,LT-β)联在一起存在于T细胞膜上。 具有促使肿瘤细胞凋亡的能力,能活化中性粒细胞、巨噬细胞、内皮细胞和B细胞等。
Interleukin-4 由活化的Th2细胞产生,作用于B细胞、T细胞、巨噬细胞、内皮细胞、肥大细胞和成纤维细胞等。刺激B细胞的生长和分化,在小鼠是诱导产生IgE的开关,在变态反应中起关键作用;促使T细胞的发育,使静止的T细胞发育成为细胞毒性T细胞,可在无IL-2的条件下使Th细胞生长。 对巨噬细胞既能提高MHC II类分子的表达、抗原提呈和细胞毒性的能力,又能降低IL-1、IL-6和TNF-α分泌。 IL-4的作用可被IFN-γ所抑制。
Interleukin-5 由Th细胞产生,促进B细胞分化与生长,诱导无活性B细胞产生IgA及活性B细胞产生IgM、IgG,诱导嗜酸性粒细胞分化及细胞毒性T细胞的增殖,从而在机体免疫调控中发挥重要作用。 Interleukin-9 只由Th2细胞产生。诱导辅助性T细胞和骨髓源性母细胞的生长,并增强IL-4对B细胞的作用。
Interleukin-10 主要由Th2细胞产生,Th0细胞、部分B细胞、巨噬细胞和角质细胞也能产生。Th1细胞、B细胞、巨噬细胞、NK细胞、母细胞和胸腺细胞是它的靶细胞,能抑制Th1细胞和NK细胞的活性,抑制巨噬细胞IL-1、IL-6和TNF-α 的分泌、MHC-II的表达及刺激IL-1RA的产生。并能抑制B细胞、胸腺细胞和母细胞的增殖。
Interleukin-13 仅由Th2细胞产生,其生物功能与IL-4类似,可诱导B细胞增殖,产生抗体,促进NK细胞产生IFN,抑制单核细胞产生炎性因子,促进单核细胞表面MHC-II类抗原的表达并增强其抗原提呈作用。 Interleukin-17 由活化的CD4+细胞产生,能促进IL-6和IL-8的产生。
趋化因子(Chemokines) 根据半胱氨酸残基的空间位置可将其分化为α和β两个亚族。 α chemokines:为多源性细胞因子,可由单核细胞、巨噬细胞、内皮细胞、角质细胞及部分T细胞产生。是中性粒细胞、T细胞及嗜碱粒细胞的趋化因子,可使中性粒细胞趋化,激活脱颗粒和嗜碱性粒细胞释放组胺,提高游走率,在炎症和免疫过程中具有重要的调节意义。
2. β chemokimes MIP-1 Marophage inflammatory protein MCP monolyte chemoattractent protein Rantes protein MIP-1由Macrophage、T、B等产生,作用于monolyte。嗜中性粒细胞、B、T细胞,其MIP-1 β吸引CD4+T,而MIP-1 α吸引B、酸性粒细胞和Tc。 MCP由M、T、成纤维细胞等细胞产生,对monocytos具化学趋向和活化作用,刺激它们破裂、释放溶酶体酶。 RANTes由T、M产生,对monolytes、M和部分T细胞具趋化作用,活化eosinophils,刺激basophics释放组胺。
抗病毒细胞因子 IFN IFN-α:lymphocyte、monocyte、M产生 IFN-β:成纤维细胞 IFN-ω:lymphocyte、monocytes、trophololast IFN-τ:反刍兽trophoblast IL-16 是一种由CD8+T细胞分泌的蛋白质,通过CD4和其它T细胞相结合,对淋巴细胞具有趋化性。
生长因子 是一大群对血细胞具分化和成熟作用的糖蛋白 1. IL-3:又称为克隆集落刺激因子(multi-CSF)。主要由Th1细胞、B细胞产生,生物学活性非常广泛,可刺激多能血干细胞和各系统细胞的分化与增殖,促进和维持肥大细胞的增殖,增强嗜酸性粒细胞的活性,促进NK细胞的杀伤活性。 2. IL-7:由骨髓细胞、脾细胞和胸腺基质细胞产生。主要作用于胸腺细胞、T细胞、B细胞、单核细胞和淋巴干细胞,诱导骨髓原B细胞、胸腺细胞和T细胞的增殖,控制淋巴干细胞的活性,促进细胞毒性T细胞的发育。
IL-11 由骨髓基质细胞和成纤维细胞产生。能刺激与IL-6相关的B细胞的生长,刺激与IL-3有关的巨噬细胞集落形成,促进肝急性反应蛋白的产生。 IL-14 由T细胞和一些恶性B细胞产生。能诱导B细胞的增殖,抑制免疫球蛋白的分泌并选择性地扩增某些B细胞亚群。
4. IL-15 由巨噬细胞、上皮细胞和成纤维细胞产生。其功能类似于IL-2,可作为T细胞和NK细胞的生长因子,刺激Th细胞和细胞毒性T细胞的增殖。 5. G-CSF MG-CSF、M-CSF及TGF-β
细胞因子的毒性效应 1. 细菌的败血性休克Bacterial septic shock