Download

1 / 56
720 likes | 1.89k Views
Estructura del Sistema Nervioso. Dra. María Mercedes Soberón L. Puntos a tratar: Estructura proteica de la membrana y moléculas de adhesión celular. Citoesqueleto de las neuronas y glias Compartimentación intra e intercelular.

E N D
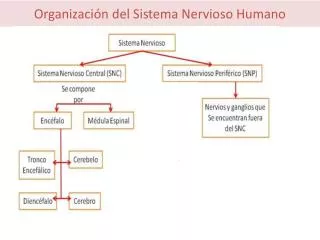
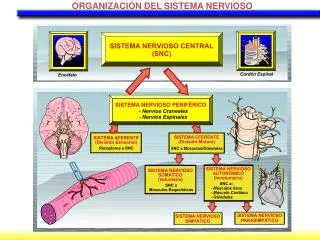
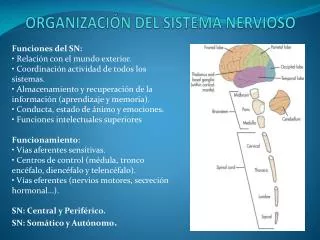
Estructura del Sistema Nervioso Dra. María Mercedes Soberón L.
Puntos a tratar: • Estructura proteica de la membrana y moléculas de adhesión celular. • Citoesqueleto de las neuronas y glias • Compartimentación intra e intercelular. • la colaboración metabólica (a través de sustratos) entre dos compartimentos. • Sustratos neuronales y gliales en la compartimentación.
¿Qué tan diferente son las membranas del sistema nervioso de las demás membranas? Espesor aprox. 8nm lo cuál la hace demasiado delgada para poder ser observada por un microscopio óptico. Es rica en canales dependientes de voltaje……
Proteínas de Membrana: Canales Iónicos Principalmente importantes en neuronas Excitabilidad neuronal depende de función de canales iónicos.
TIPOS DE CANALES IÓNICOS • Abiertos por voltaje. Conformación depende de diferencia de carga iónica a ambos lados de membrana. • 2. Abiertos por ligandos. Conformación depende de unión con ligando (neurotrasmisores, nucleótidos) que no es el soluto que pasa por el canal)
Estructura General de un Canal Iónico • Un túnel central o poro a través del cual fluyen iones con respecto a su gradiente electroquímico. • 2) Un filtro de selectividad, el cual dicta a cuál ión se le permitirá el paso a través del poro, denominado región P. • 3) Una estructura que ejerce la función de compuerta controlando la probabilidad conformacional de apertura y cierre del canal Filtro Memb. celular Memb. celular Compuerta
Canal de Potasio Bacteriano (KcsA) Compuestos por 4 subunidades C/subunidad: M1 y M2 Filtro selectividad recubierto por pentapéptido: Gly-Tyr-Gly-Val-Thre. Grupos C=O de pentapéptido forma recubrimiento del filtro Filtro Memb. celular Memb. celular M2 M1 Compuerta
Canal iónico de Potasio activados por voltaje (Kv) en Eucariotas, se abren por cambios en el voltaje Presentan dos dominios importantes: • DOM. PORO: • 4 subunidades homólogas dispuestas simétricamente con hélices S5 y S6 conformando la “compuerta” del poro • Filtro selectivo que promueve paso de K+
2. DOM. SENSOR DE VOLTAJE: Hélices S1 – S4. S4 elemento clave del sensor de voltaje. Cambio de potencial más positivo, produce movimiento de S4 que tira de S6 y abre compuerta ¿Cómo se desactiva el canal? • Porción N-terminal de péptido de desactivación se ajusta dentro de cavidad citoplasmática del poro • Desactivado, liberación de péptido y cierre de compuerta Péptido de desactivación
canales de potasio dependientes de voltaje KCNQ/(tipo M) ejercen un importante control sobre la excitabilidad neuronal.
Se han identificado mutaciones en los cuatro miembros conocidos de la familia KCNQ que son responsables de ciertas formas hereditarias de epilepsia, de arritmia y de sordera en humanos. CAUSAS 1. Proteínas que lo forman no se ensamblen correctamente y, por tanto, el canal queda retenido en el interior de las neuronas y no puede alcanzar la superficie celular. Despolarización -50 mV
Canales de Potasio 1) Canales de K +dependientes de voltaje (Kv) 2) Canales de potasio rectificadores hacia adentro (Kir) Permiten el paso del potasio hacia el intracelular cuando la membrana plasmática se encuentra a un voltaje más negativo que su potencial de equilibrio y se encuentran cerrados a potenciales más positivos
3) Estructura molecular de un canal de K+ sensible a ATP receptor de sulfonilurea (SUR ) regula la apertura o cierre del poro (Mg-ADP lo activa) (inhibe el canal) Cada una de estas familias de canales de potasio puede a su vez presentar subfamilias, dependiendo de la electrofisiología del canal o del ligando que medie la apertura de tales proteínas
Canal Ca++ operado por voltaje Subunidad . 2 La subunidad 2 extracelular se une a la transmembrana mediante un puente disulfuro. Sub unidad .1Estructura transmembrana helicoidal que consta de 4 dominios repetidos que rodean el poro. subunidad consta de 4 dominios transmembrana Subunidad . sirve de anclaje para la subunidad .1
Canal Ca++ operado por voltaje Tipo N: Canales Cav2 Constituido por canales de tipo N que corresponden a la subunidad 1, se encuentran presentes en el tejido nervioso y tienen 3 variantes : Cav2.1, Cav2.2 y Cav2.3. Mutaciones del gen que codifica la subunidad Cav2.1 están asociadas a enfermedades degenerativas del cerebelo y ataxias.
Defectos en canales iónicos que son causa de enfermedades hereditarias Mayoría de trastornos afectan movimiento de iones a través de membrana de cél. musculares, nerviosas y sensoriales lo que reduce la capacidad de estas células para desarrollar o trasmitir impulsos
Receptores de Membrana • Tienen dos componentes importantes: • Componente de fijación, que protruye al exterior de la membrana en el surco sináptico, y fija el neurotransmisor liberado de la terminal presináptica. • Componente ionóforo, que penetra desde la membrana al interior de la neurona
El componente ionóforo puede ser de 2 tipos: - Un conducto de iones activado químicamente - Conductos activados por ligando, cuyo paradigma es el NMDA (-N-metil-D-aspartato-), verdadero conglomerado o complejo de canales iónicos, que pueden ser de 3 tipos principales: Canales de sodio, Canales de potasio y Canales de cloruro. También pertenecen a esta categoría los receptores denominados AMPA (ácido propionico de alfa amino 3 hidroxi 5 metil 4 isoxazol) /Kainate
Tipos Receptores de Membrana 1. Ionotróficos 2. Metabotróficos
Receptores Ionotróficos Determinan la apertura o cierre de canales debido a que producen despolarizaciones o hiperpolarizaciones. Es una respuesta rápida. Algunos canales pueden necesitar la unión de dos neurotransmisores como es el caso del receptor de Acetilcolina o el receptor NMDA que necesita glutamato y glicina. La señalización tambien puede ser intracelular, generalmente fosforilando en la cara citoplasmica del canal el receptor, induciendo la apertura del canal.
Receptores ionotrópicos de los aminoácidos excitatorios • En razón de su afinidad por agonistas sintéticos: NMDA, AMPA y Kainato. • la familia del receptor NMDA(-N-metil-D-aspartato), • la familia de los receptores AMPA (ácido propionico de alfa amino 3 hidroxi 5 metil 4 isoxazol) • La familia de receptores KAINATO Estudios de clonaje de receptores han mostrado que hay un largo número de potenciales subtipos de receptores en ambas familias.
Compuestos de subunidades NR1 y NR2. Son permeables al influjo de Ca2+ y al flujo retrógrado de K+. El sitio de unión del glutamato está localizado en la subunidad NR2, mientras que el sitio de unión de su coagonista obligatorio glicinaestá situado en la subunidad NR1. Estructura de Receptores NMDA
Cuando se abre el canal regulado por los NMDA, el Ca+2 y el Na+ entran en la neurona y el K+ lo abandona, dependiendo del potencial de la membrana postsináptica. Glutamato sólo provoca la abertura del canal y cuando se une al receptor NMDA, el poro se abre. Sin embargo, con potenciales de membrana en reposo negativos normales, el canal se bloquea por lones Mg+2y el “bloqueo de magnesio” impide que otros iones entren libremente en la célula. El Mg+2 sólo sale del poro cuando la membrana está desporalizada, lo que habitualmente ocurre tras la activación de los canales de AMPA en las mismas sinapsis y en las vecinas.
Compuesto de cuatro subunidades, c/subunidad compuesta a la vez de 3 dominios transmembrana y una solo dominio intra citoplasmático. Pueden ser receptores homomérico, es decir tener subunidad del mismo tipo (ya sea GluR1, GluR2, GluR3 o GluR4), o como un heterómero conformado por 4 subunidades distintas. Permite la entrada mayormente de Na+ y sale K+, entrando sólo determinada cantidad de Ca++ en el grupo de la subfamilia GluR2. Receptor AMPA moduladores positivos : ciclotiazida y aniracetam. Antagonistas: NBQX, GYKI y zinc.
Estructura del receptor GABA-B Se encuentra en la membrana plasmática tanto del terminal presináptico como del terminal postsináptico. No está emparentado con canales de cloro como el receptor GABA-A, sino que modulan canales de calcio y de potasio por una interacción con la proteina G y la adenil ciclasa.
Receptor Nicotínico Receptor de 250-270 kDa es un pentámero formado por cuatro tipos de subunidades distintas, 2α, β, , , total 5 subunidades. Las cinco subunidades se encuentran ordenadas alrededor del eje central del poro, que se encuentra formado por las hélices M2 de cada subunidad presentando la estructura una simetría pentagonal.
Receptor Nicotínico Anillo de residuos hidrofóbicos de Leu de hélices M2, se encuentran tan próximas unas a otras que producen cierre del canal, impidiendo el movimiento de iones. Unión de acetil colina a subunidades , induce cambios conformacionales alostericos originando ligero giro de hélices, que mueve estas cadenas laterales hidrofóbicas alejándolas del centro del canal y permitiendo el paso de iones. cerrado abierto
Canales iónicos operados por vías metabólicas activadas por proteína G El neurotransmisor actúa sobre el receptor, que activa una proteína G, que activa la adenil ciclasa y que transforma el ATP en AMPcíclico. Este AMPcíclico puede actuar sobre el canal de membrana, que al fosforilarlo, se abre.
Liberan mensajeros intracelulares (AMPcíclico, DAG y fosfolípidos). Cuando el receptor recibe el neurotransmisor, pone en funcionamiento la adenilatociclasa y el ATP se transforma en AMPcíclico. Una vez actuado, es destruido por la fosfodiesterasa. EL AMPcíclico activa una proteinquinasa, que fosforila una proteína. Receptores Metabotróficos Las neuronas granulares de cerebelo en cultivo presentan receptores metabotrópicos de nucleótidos de tipo P2Y6, cuyo agonista fisiológico específico es el nucleótido, uridina difosfato, UDP
Molécula señalizadora Receptor unido a Proteína-G Fosfolípido inositol Fosfolipasa C activada Proteinquinasa C activada Subunidad -Prot-G activada Lumen del retículo endoplásmico Las Dos Ramificaciones de la Ruta de Fosfatidil Inositol
Los receptores de Ca+2, cuando reciben el neurotransmisor, abren 1 canal de Ca+2, entra Ca+2 extracelular y se junta a la proteína calmodulina, formando la calmodulina-Ca,++ que activa una proteinquinasa que fosforila una proteína. Complejo Ca++-calmodulina
Receptor Muscarínico El receptor muscarínico tiene forma de serpentina acoplado a una proteína G, adenilciclasa y fosfolipasa Es el tipo predominante de receptor colinérgico en el cerebro, donde parecen hallarse involucrados en la memoria y aprendizaje (pueden estar involucrados en trastornos como la depresión y manía). Los receptores muscarínicos superan a los nicotínicos en un factor de diez a cien.
Proteínas específicas presente en SN • La GP-350 una sialoglicoproteína, soluble unida a la membrana, es específica del cerebro y está localizada en las células piramidales y estrelladas. • La sinaptina contenida en las vesículas sinápticas y en las membranas plasmáticas de la sinapsis • La D1, D2 y D3 son proteínas específicas del cerebro, localizadas en las membranas sinápticas y que difieren en su peso molecular. • La P-400, proteína que está unida a las membranas y que se halla solamente en el cerebelo, en las dendritas de las células de Purkinje.
Proteína que pareciera ser importante para ayudar al crecimiento y la supervivencia de las neuronas. La APP puede ayudar a curar a aquellas neuronas lesionadas y a que algunas partes de las neuronas se desarrollen después de una lesión cerebral. Proteína Precursora de Amiloidea (APP) Está asociada con la membrana celular. Una vez que se ha formado, la APP penetra la membrana de la neurona y queda parcialmente adentro y parcialmente afuera de la célula.
Función del Citoesqueleto • Mantenimiento forma asimétrica de las neuronas. • Concentración diferentes elementos estructurales en sitios específicos del citoplasma, membrana o núcleo. • Establecimiento de conexiones interneurales y formación de sinapsis
Experimentos con animales en estados de depresión han demostrado alteraciones en el citoesqueleto: significativa reducción en subunidad ligera de neurofilamentos (Reines A. y col. Neuroscience 2004, 129:529-538)
Componentes del Citoesqueleto microtúbulo microfilamento Filamento intermedio o neurofilamento
Microtúbulos, microfilamentos, proteína unida a microtúbulos 2 (MAP2) Microtúbulos, Neurofilamentos, Proteína TAU
¿Cómo es que las vesículas y los cargamentos sin membrana pueden moverse a lo largo del citoesqueleto? Proteínas motoras generan fuerzas necesarias para mover objetos dentro de una célula - Cinesinas y Dineínas se mueven a lo largo de los microtúbulos. - Miosinas se mueven a lo largo de microfilamentos
Se unen al microtúbuloy actúan como máquinas generadoras de fuerza que hidrolizan ATP. Utilizan al microtúbulo como riel Cabezas globulares Cinesina • Tetrámero: • - Dos cadenas pesadas idénticas • Dos cadenas ligeras idénticas • Transporte de organelas mediado por cinesina + - dominio cola reconocimiento de membrana lipídica de vesículas a transportar, probablemente por mediación de las cadenas ligeras.
Dineína Cabeza generadora de fuerza Purificada de tejido cerebral. Consta: dos cadenas pesadas y varias cadena intermedias y ligeras. Se mueve de manera progresiva a lo largo del microtúbulo hacia el extremo MENOS del polímero, contrario al sentido de la cinesina Dineína Dineína - + Cinesina
Sinapsis en SNC Sitios de contacto Célula-Célula altamente especializados • Proteínas transmembranas de adhesión celular resultan ser importantes en el proceso de sinapsis: • cadherinas, • integrinas, • neurexinas-neuroliginas, • superfamilia de Inmunoglobulinas, etc.
Moléculas de Adhesión Celular (CAM) Glicoproteínas de membrana que participan en la adhesión celular. Ligandos Extremocarboxilo terminal, se encuentra fijo en citoplasma y en citoesqueleto. La región transmembrana, atraviesa la membrana celular. Receptores Membrana plasmática Resto de glicoproteína se ubicaextracelularmente y termina en un grupo amino.El aminoterminal, da especificidad a la molécula para unirse a otras CAMs. Citoplasma COOH
Función Se enlazan a receptores situados sobre la superficie de las células. • Estructuralmente existen cuatro familias de CAMS: • Cadherinas • Integrinas • Selectinas • 4. Superfamilia inmunoglobulinas Dependientes de Ca++