Download

1 / 76
770 likes | 1.09k Views
古生菌. Archaea. 主讲人:杨智越 徐德阳. 西伯利亚细菌. 苏丝曼拍摄的最为古老的活生物是俄罗斯西伯利亚地区的细菌 Actinobacteria 。这些真正意义上的“老寿星”是在一个冰芯中发现的。 DNA分析结果显视该古菌的年龄在40万岁到60万岁之间。苏丝曼曾前往丹麦哥本哈根的尼尔斯·波尔研究所,利用显微镜给 Actinobacteria 拍照。. 目录. 第一节 古生菌的命名与发现由来 第二节 古生菌的形态与结构特征 第三节 古生菌的类群 第四节 古生菌的应用

E N D
古生菌 Archaea 主讲人:杨智越 徐德阳
苏丝曼拍摄的最为古老的活生物是俄罗斯西伯利亚地区的细菌Actinobacteria。这些真正意义上的“老寿星”是在一个冰芯中发现的。 DNA分析结果显视该古菌的年龄在40万岁到60万岁之间。苏丝曼曾前往丹麦哥本哈根的尼尔斯·波尔研究所,利用显微镜给Actinobacteria拍照。
目录 第一节 古生菌的命名与发现由来 第二节 古生菌的形态与结构特征 第三节 古生菌的类群 第四节 古生菌的应用 第五节 古生菌的研究意义
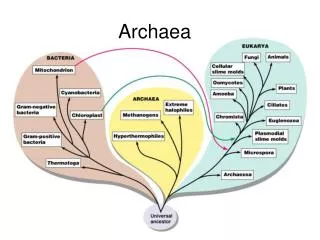
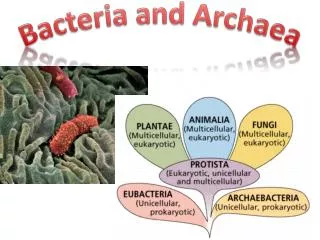
第一节 古生菌的命名与发现由来 古生菌,又名古细菌或古核生物是一些生长在极端特殊环境中的细菌,这些特殊环境包括大洋底部的高压热溢口、热泉、盐碱湖等。
古细菌这个概念是1977年由Carl Woese和George Fox提出的,原因是它们在16SrRNA的系统发生树上和其它原核生物有所区别。这两组原核生物起初被定为古细菌和真细菌两个界或亚界。而Woese认为它们是两支根本不同的生物,通过对不同细菌的rRNA样品进行检测分析,他重新将其命名为古菌和细菌,这两支和真核生物一起构成了生物的三域系统。
第二节 古生菌的形态与结构特征 一、古生菌的形态 古生菌的细胞形态有球形、杆状、螺旋形、耳垂形、盘状、不规则形状、多形态,有的很薄、扁平,有的有精确的方角和垂直的边构成直角几何形态,有的以单个细胞存在,有的呈丝状体或团聚体。其直径大小一般在0.1~15μm,丝状体长度有200μm。
二、古生菌的结构 古生菌细胞有细胞质、细胞膜和细胞壁三种结构。古生菌细胞也有一层把细胞和外部环境隔离开的外膜。在膜内包裹着细胞质,其中悬浮着DNA,古生菌的生命活动在这里进行。几乎所有的古生菌细胞的外面都围有细胞壁,这是一层半固态的物质,它可以维持细胞的形状,并保持细胞内外的化学物质平衡。
三、古生菌与其它生物的最显著的化学差异 古生菌与其它生物最显著的化学差异是体现在生物膜上,其主要差异有四点: (1)甘油的立体构型。组成细胞膜的基本单位是磷脂。用来构建古生菌磷脂的甘油是细菌和真核生物细胞膜上所用的甘油的立体异构体。细菌和真核生物的细胞膜中是D型甘油,而古生菌的是L型。
(2)醚键。当侧链加到甘油分子上时,大多数生物是通过酯键来结合的,即加上两个氧原子而连接到甘油的一端。其中一个氧原子用来与甘油形成化学键,另一个原子则在发生结合后伸出来。相反,古生菌的的侧链是通过醚键来连接的,没有伸出来的氧原子。因此,古生菌的磷脂的化学性质不同于其它生物的膜脂。
(3)类异戊二烯链。细菌和真核生物的磷脂上的侧链通常是链长16到18个碳原子的脂肪酸,古生菌膜上的磷脂则不是脂肪酸,而是由异戊二烯构成的由20个碳原子形成的侧链。 细菌细胞膜结构 古菌酯类单层结构
(4)侧链的分支。古生菌细胞膜的侧链由不同的化合物构成,细菌和真核生物的脂肪酸没有这些侧分支。侧分支能够形成碳原子环。这种环可以稳定膜上的结构,有助于古生菌生活在高温中。
古菌之所以能够存在于高温、 高盐以及强酸性等极端条件下,得益于它特殊的侧链及成键的分子特性.古菌的侧链多为不含活性官能团的碳长链分子,与甘油酸分子以非常稳定的醚键相联,并且组成细胞膜的脂类为一整个长碳链分子,形成单层细胞膜. 这种相当稳定的结构使得古菌细胞膜非常坚固,能够抵抗即使是非常恶劣的外界条件.
第三节 古生菌的类群 古生菌的主要类群有以下四种: (1)产甲烷菌 (2)极端嗜盐菌 (3)极端嗜热菌 (4)嗜热嗜酸菌
一、产甲烷菌 1.1 产甲烷菌的形态结构 产甲烷细菌是一群迄今为止所知的最严格厌氧的能形成甲烷的化能自养或化能异养的古菌群。它们的形态以杆状和长丝状为主。细胞的宽为0.5-1.0um,不形成芽孢,产生鞭毛,不运动。革兰氏染色反应不定。
1.2 产甲烷菌的分类 随着厌氧培养技术和菌种鉴定技术的不断成熟,产甲烷菌的系统分类也在不断完善. 《伯杰系统细菌学手册》第9版将近年来的研究成果进行了总结和肯定,并建立了以系统发育为主的产甲烷菌最新分类系统. 产甲烷菌分可为5个大目,分别是:甲烷杆菌目(Methanobacteriales)、甲烷球菌目(Methanococcales)、甲烷微菌(Methanomicrobiales)、甲烷八叠球菌目(Methanosarcinales)和甲烷火菌目(Methanopyrales) 。
1.3 产甲烷菌的化学组成 在化学组成上与其他细菌差距甚大。它的细胞壁不含有真正的肽聚糖,而是含有假肽聚糖。细胞壁没有细菌细胞壁胞壁质的特征,而是富含有各种表层蛋白。其细胞膜的主要成分为植烷醇基甘油醚。
1.4 产甲烷菌的生活环境 产甲烷菌主要生活在多水、缺氧的沼泽和湿地等环境中。它们通过将CO2还原成CH4而获得能量生长,电子供体仅限于H2、甲酸和CO。氨为唯一氮源,硫化物可作为硫的来源。
二、极端嗜盐菌 2.1 极端嗜盐菌的形态 极端嗜盐菌是古细菌的典型代表之一。它们的细胞常呈现杆状、球状、三角形、多角形、盘形等多种形态。革兰氏染色呈阴性。极生鞭毛,运动或不运动。好氧或兼性厌氧。细胞内含有各种色素。
这种细菌是英国微生物学家安东尼·瓦尔斯比于1980年发现的,地点在红海附近一个盐分含量很高的水池里。它身长约0.15微米,呈方形,样子有点像邮票,这在细菌中极为罕见。这种细菌是英国微生物学家安东尼·瓦尔斯比于1980年发现的,地点在红海附近一个盐分含量很高的水池里。它身长约0.15微米,呈方形,样子有点像邮票,这在细菌中极为罕见。
2.2 极端嗜盐菌的化学组成及生活环境 细胞壁也不含有肽聚糖。细胞膜成分为类异戊二烯甘油醚酯。它们具有利用光能驱动质子泵的作用,因此可利用质子梯度所产生的能量合成ATP。极端嗜盐菌主要分布在高盐环境中,如盐湖、盐碱湖、晒盐场、以及含盐量高的土壤中等。
2.3 极端嗜盐菌的嗜盐机制 2.3.1 嗜盐菌的Na+依存性 (1)Na+与细胞膜成分发生特异作用而增强了膜的机械强度,有利于细胞膜结构的稳定。 (2)在细胞膜的功能方面,嗜盐菌中氨基酸和糖的能动运输系统内必需有Na+存在, 而且Na+作为产能的呼吸反应中一个必需因子起着作用。 (3)Na+被束缚在嗜盐菌细胞壁的外表面,起着维持细胞完整性的重要作用。
2.3.2 嗜盐菌中酶的盐适应特性 嗜盐酶只有在高盐浓度下才具有活性,盐去除后,嗜盐酶就会失活。 由海洋细菌中分离的酶,就它们与盐的依存关系可分为三类:
(1)为不加盐时,酶活性最高,加盐就受抑制。在这类嗜盐菌中可能存在某种保护机制。(1)为不加盐时,酶活性最高,加盐就受抑制。在这类嗜盐菌中可能存在某种保护机制。 (2)为不加盐时有一定活性,加盐时酶活力进一步增强,最适盐浓度低于细胞内离子浓度,过高浓度的盐会使酶活性受抑制。 (3)为不加盐时几乎不显示活性。由于盐的作用使酶强烈的活性化。
2.3.3 嗜盐菌质膜、 色素及 H+泵作用 (1)嗜盐菌质膜、 色素 嗜盐菌具有异常的膜。嗜盐菌细胞膜外有一个亚基呈六角形排列的 s单层,这个所谓的“s 单层”由磺化的糖蛋白组成,由磺酸基团的存在使 s 单层呈负电性,因此使组成亚基的糖蛋白得到屏蔽,在高盐环境中保持稳定。
低氧压或厌氧情况下光照养, 盐生盐杆菌(H . halobium)产生红紫色菌体, 这种菌体的细胞膜上, 有紫膜膜片组织, 约占全膜的 50%, 由 25%的脂类和75%的蛋白质组成。目前,对盐生盐杆菌的细胞视紫红质研究最透彻, 由三个细胞视紫红质分子构成的三聚体可在细胞膜上形成一个刚性的二维六边形的稳定特征结构,即紫膜。
(2)嗜盐菌H+泵作用 嗜盐菌的菌视紫素可强烈吸收 570nm 处的绿色光谱区,菌视紫素的视觉色基(发色团) 通常以一种全—反式结构存在于膜内侧。 当受光照时,分子中的生色团(一视黄醛)在光刺激下产生快速的从全反结构到 13 顺结构的光异构化过程。
这种转型作用的结果使H+质子经转移到膜的外面,然后通过热弛豫,历经一系列中间态再回到基态 B,完成顺式状态又转换成更为稳定的全—反式异构体的一次光循环。 这种循环,形成质膜上的H+质子梯度差,即H+泵,产生电化势,菌体利用这种电化势在ATP 酶的催化下,进行ATP的合成,为菌体贮备生命活动所需要的能量。
2.3.4 排盐作用 嗜盐菌的生长虽然需要高钠的环境,但细胞内的Na+浓度并不高,如盐杆菌光介导的H+质子泵具有Na+/K+反向转运功能,即具有吸收和浓缩K+和向胞外排放Na+的能力。K+作为一种相容性溶质,可以调节渗透压达到细胞内外平衡,其浓度高达 7mol/ L ,以维持内外同样的水活度。
2.3.5 嗜盐细胞内溶质浓度的调节 嗜盐微生物由于产生大量的内溶质或保留从外部取得的溶质而得以在高盐环境中生存。 氨基酸在嗜盐细胞内溶质浓度调节中起着重要作用。随培养基食盐的增加,氨基酸浓度有规律的增加,其中主要是谷氨酸和脯氨酸,及甘氨酸,它们具有渗透保护作用,是溶质浓度调节的重要因子。
2.3.6 耐盐基因片段 目前,已相继分离和鉴定了与渗透保护物合成途径有关的结构基因,如proABC和bet 基因,以及转录表达受到渗透压调控的一些操纵子,如kdp 操纵子, pro和opu系统以及作为全细胞调节因素的 ompR系统, 并且进行了有关耐盐基因的分离和转移研究。
有关嗜盐菌遗传学方面的研究国外已有大量报道,但由于这类菌难以得到合适的选择标记,突变株不稳定等特性,进展缓慢,目前在嗜盐菌中发现了质粒,但这些质粒的分子量较大,拷贝数低,缺乏便于克隆外源DNA 的单一酶切位点,而不能用作载体。
三、极端嗜热菌 3.1 极端嗜热菌简介 极端嗜热菌是一类偏喜高温的古生菌。他们中的所有成员都具有80℃以上的最适温度。绝大多数为专性厌氧菌。营养类型有化能有机营养型、化能无机营养型以及兼性营养型。
3.2 极端嗜热菌的生活环境 极端嗜热菌广泛分布于温泉、肥堆、煤堆、有机物堆、强烈太阳辐射加热的地面、地热区土壤以及陆地和海底火山口等高温环境。在冰岛, 有一种嗜热菌可在98℃的温泉中生长。在美国黄石国家公园的含硫热泉中, 曾经分离到一株嗜热的兼性自养细菌——酸热硫化叶菌(Sulfolobus), 可在高于90℃温度下生长。
3.3 极端嗜热菌的耐热机制 嗜热微生物能够在高温环境中生长并繁殖,有其独特的适应机制,特殊结构的细胞膜、蛋白质、遗传物质都与之有关。 (1)特殊的细胞膜 具体内容参见第一节
(2)稳定的天然结构 蛋白质的一级结构中,个别氨基酸的改变会影响整体的热稳定性。嗜热微生物中蛋白质和酶的一级结构与常温菌同源蛋白相比,氨基酸的组成和顺序有明显的差异,具有高的热稳定性。卢柏松等人发现嗜热菌蛋白质中Leu,Pro,Glu和Arg的含量高于常温菌,这些氨基酸都有利于提高蛋白质的稳定性。
(3)促进热稳定的因素 细胞内促进蛋白质热稳定的因素,主要是金属离子和一些特殊的蛋白质。金属离子的化学修饰作用对提高蛋白质热稳定性有很大贡献。例如,Ca2 +、Mg2 +、Zn2 +等都能起到稳定蛋白质空间结构的作用。嗜热微生物在高温环境中能产生一些特殊蛋白质,为嗜热菌上限高温生长时所必须。
(4)DNA结构的热稳定性 大多数嗜热微生物生长上限温度和G+C%之间存在正相关关系,嗜热菌DNA结构中G+C含量高,通常为53.2%。DNA双螺旋结构的稳定性主要是由碱基堆积力和氢键来维持,DNA中G+C%含量越高,氢键数量就越多,碱基堆积力就越大,解链所需的温度就越高,DNA双螺旋结构就越稳定。
嗜热菌的DNA双螺旋结构中核苷酸排列非常有序,很少有突出的核苷酸或其他不规则因素的存在,同时嗜热菌DNA双螺旋结构中螺距较常温菌短,有利于DNA的稳定。
(5)tRNA的热稳定性 嗜热菌tRNA中G+C%含量高,有很好的热稳定性,并且嗜热菌的tRNA周转速度快,能够保护热不稳定的代谢物,使重要代谢产物迅速合成。极端嗜热菌tRNA转录后经过相关的修饰有助于茎环上碱基之间的相互作用、D-环的堆积和新的氢键及离子对的产生,有利于适应高温环境。
同时,嗜热菌核糖体的热稳定性是嗜热菌生长上限温度的决定性因子。此外,RNA螺旋侧面突起的碱基和近端酯键之间的配对能避免结构上的不稳定性。
四、嗜热嗜酸菌 嗜热嗜酸菌一般是没有细胞壁的细菌。它们专性嗜热嗜酸且只在极低pH条件下才能生长。它们形态具有多样性。仅有一个三层结构的细胞膜,无真正的细胞壁,膜中的脂质为植烷基二甘油四醚。革兰氏染色呈阴性。营养类型为兼性厌氧型。它们往往生长在火山区或含硫量极为丰富的地区。能氧化还原态的硫化物和金属硫化物,以从其中取得能量。
当然,古细菌并不是仅局限于这四大类。研究还发现有些古生菌为极端嗜碱菌、极端嗜压菌、极端嗜冷菌等等。这些不同类型的古生菌各自有着不同的形态特征、生理功能及生活条件。对于这些古生菌的研究有着重要的生物学意义。尤其是极端微生物代谢产生特有的生物活性物质以及自身的一些活性物质,已广泛应用于许多领域。
第四节 古生菌的应用 一、产甲烷菌的应用 1.1 在废水处理方面的应用 嗜冷产甲烷菌对于自然界的碳素循环具有非常重要的意义,近年来引起了国内外学者的广泛关注。利用嗜冷产甲烷菌实现低温厌氧生物处理过程,可从本质上突破低温厌氧工艺的技术瓶颈,进而大大拓展厌氧生物处理技术的应用范围并降低废水处理的成本。
1.2 在酿酒工业上的应用 泸州老窖古酿酒窖池中,多种厌氧细菌类群参与曲酒发酵生香,其中己酸菌具有特别重要的生产性能. 上世纪80年代首次从泸州老窖泥中分离出氢营养型的布氏甲烷杆菌CS菌株,揭示了酿酒窖池是产甲烷古菌存在的又一生态系统。 随后发现该菌和从老窖泥中分离的己酸菌——泸酒梭菌菌株存在“种间氢转移”互营共生关系,混合培养时可较大程度提高己酸产量。
以后将CS菌株应用于酿酒工业,与己酸菌共同促进新窖老熟,有效提高酒质。 因此,窖泥中栖息的产甲烷古菌既是生香功能菌,又是标志老窖生产性能的指示菌。在对泸州古酿酒窖池微生物研究中,观察到窖泥中的产甲烷古菌在形态特征、营养类群和生理特性上表现出多样性,并分离到部份菌株。
1.3 在煤层气开发中的应用 生物成因煤层气是在较低的温度条件下,有机质通过各种不同类群细菌的参与或作用,在煤层中生成的以甲烷为主的气体。产甲烷菌对煤层气的形成起着重要的作用,目前已发现产甲烷菌有低温型、中温型和嗜热型。