Download
1 / 17
170 likes | 293 Views
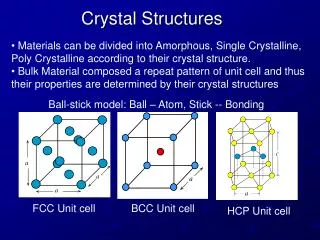
Free Energy of Transfer for Protein Burial Crystal structures of 37 proteins were examined amino acid surface amino acid buried f = (N b / N b )/(N s /N s ) where N b = frequency of amino acid burial in the protein interior

E N D
Free Energy of Transfer for Protein Burial Crystal structures of 37 proteins were examined amino acid surface amino acid buried f = (Nb/Nb)/(Ns/Ns) where Nb = frequency of amino acid burial in the protein interior Ns = frequency of occupancy on the protein surface by the same amino acid Nb;Ns : sum of amino acids buried; on surface burialG = -RTln f Criterion used for surface/burial: accessable surface area ASA ASA = surface area of aminoacid in the protein area of aminoacid in the extended tripeptide glyNHRCHCOgly where R = amino acid side chain in question Miller, S.; Janin,J.; Lesk, A. M. Chothia. J. Mol. Biol 1987. 196, 641.
Side Chain Hydrophobicity Solubility of R-CHCONH2 in water octanol | NHCOCHwhere R = amino acid side chain K = solubility in 1-octanol solubility in water woG = -RTln K Fauchere, L.; Pliska, V. Eur J.Med. Chem.-Chim. Ther. 1983, 18, 369.
Free Energy of Tranfer For Residue Burial (DburialG) and Hydrophobicity (DwoG) at 300 K. R DwoG DburialG kJ mol-1 alanine -CH3 -1.8 -0.8 asparagine -CH2CONH2 3.4 2.9 aspartic acid -CH2CO2H 4.4 3 cysteine -CH2SH -5.6 -2.8 glutamine -CH2CH2CONH2 1.3 3.1 glutamic acid -CH2CH2CO2H 3.6 4.6 histidine -CH2(C3H3N2) -0.8 -0.2 isoleucine -C(CH3)CH2CH3 -10.3 -3.1 leucine -CH2CH(CH3)2 -9.7 -2.7 lysine -(CH2)4NH2 5.6 8.4 methionine -CH2CH2SCH3 -7 -3 phenylalanine -CH2C6H5 -10.2 -2.8 serine -CH2OH 0.2 1.4 threonine -CH(CH3)OH -1.5 1.1 tryptophane -CH2(C8H6N) -12.8 -1.9 tyrosine -CH2(C6H4OH) -5.5 0.9 valine -CH(CH3)2 -6.9 -2.6
Why is there scatter in the plot of woG vs burialG ? Consider an aminoacid in the exterior of a protein and compare it to an aminoacid in the interior. Then consider the same aminoacid in water and in octanol.
Is there anyway to correct for loss of side chain mobility on the interior of the protein?
Contributions to Entropy Accompanying Melting Sfus = Strans + Srot + Svib + Selec; Srot = Srigidbody rot + Sconf During melting, let’s assume that Svib + Selec are not significantly affected. For an ideal gas at 298 K: Strans = 37.0 + 3/2Rln(M/40); Srigidbody rot = 11.5 + R/2ln(Im3/e) + Rln(n) where: M = molecular weight; Im3 = product of the three moments of inertia about the center of gravity of the molecule; e = external symmetry number; n = number of optical isomers. Since translation and rigid body molecular rotation vary as the logarithm of molecular weight and moment of inertia, respectively, their contribution to the total entropy change would be similar for a related group of compounds.
If the contribution of an amino acid side chain (R) to the entropy of fusion is compared to that of a standard (CH3), then Sfus(R) – Sfus(CH3) = Strans + Srigid body rot + Sconf and if: Strans(R) Strans(CH3); Srigid body rot(R) Srigid body rot(CH3); then: Sfus(R) – Sfus(CH3) Sconf or Sconf(R) where Sconf(R) is the entropy change as a result of the gain or loss of conformational flexibility relative to methyl.
R DtpceS TDDS DwoG DwoG DburialG -TDDS alanine -CH3 17.6 0 -1.8 -1.8 -0.8 asparagine -CH2CONH2 35 5.1 3.4 8.7 2.9 aspartic acid -CH2CO2H 37.3 5.8 4.4 10.2 3 cysteine -CH2SH 30.1 3.7 -5.6 -1.9 -2.8 glutamine-CH2CH2CONH2 42.1 7.2 1.3 8.5 3.1 glutamic acid -CH2CH2CO2H 44.3 7.9 3.6 11.5 4.6 histidine -CH2(C3H3N2) 32.2 6.0 -0.8 5.2 -0.2 isoleucine -CH(CH3)CH2CH3 25.9 2.4 -10.3 -7.9 -3.1 leucine -CH2CH(CH3)2 25.9 2.4 -9.7 -7.3 -2.7 lysine -(CH2)4NH2 76.2 17.3 5.6 22.9 8.4 methionine -CH2CH2SCH3 33.9 4.4 -7 -2.6 -3 phenylalanine -CH2C6H5 34.5 5.0 -10.2 -5.2 -2.8 serine -CH2OH 23.6 1.8 0.2 2 1.4 threonine -CH(CH3)OH 24.2 2.0 -1.5 0.5 1.1 tryptophane -CH2(C8H6N) 43.5 7.6 -12.8 -5.2 -1.9 tyrosine -CH2(C6H4OH) 39.9 6.6 -5.5 1.1 0.9 valine -CH(CH3)2 18.8 0.4 -6.9 -6.5 -2.6
“Protein side chain conformational entropy derived from fusion data - comparison with other empirical scales” Sternberg, M. J. E.; Chickos, J. S. Protein Engineering 1994, 7, 149 - 155.