Download

1 / 32
360 likes | 793 Views
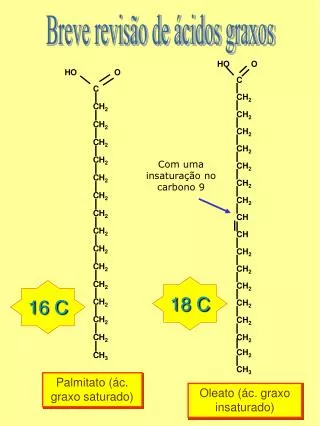
Síntese e Degradação de ácidos graxos. A pedra fundamental da síntese é o acetil-CoA , o qual é carboxilado à malonil-CoA. A carboxilação é dependente de ATP O CO 2 é posteriormente perdido em uma reação de condensação.

E N D
A pedra fundamental da síntese é o acetil-CoA, o qual é carboxilado à malonil-CoA. A carboxilação é dependente de ATP O CO2 é posteriormente perdido em uma reação de condensação. A descarboxilação espontânea será direciona a reação de condensação
Acetil CoA Carboxilase HCO3- + ATP + acetil-CoA ADP + Pi + malonil-CoA
Acetil-CoA Carboxilase catalisa a reção em 2 passos pela qual a acetil CoA é carboxilaso à malonil-CoA. Como em outras reações de carboxilação, o grupo prostético é a biotina.
Acetil-CoA Carboxilase é a enzima chave na síntese de AG A enzima de mamíferos é regulada por • fosforilação • Controle alostérico por metabólicos locais. • Mudanças conformacionais associadas à regulaçao: • Na conformação ativa, Acetil-CoA Carboxilase associa-se para formar um complexo filamentoso, multimérico; • Na conformação inativa há dissociação rendendo os monômeros da enzima.
Quinase ativada por AMP catalisa a fosforilação da Acetil-CoA Carboxilase, causando inibição. A produção reduzida demalonil-CoA previne a síntese de AG quando os estoques celulares de energia estão depletados. (AMP alto e ATP baixo).
Quinases ativadas por AMP tem um papel significante em muitos tecidos como músculo cardíaco, que não sintetizam AG Nestes tecidos, malonil-CoA, produzido via uma isoforma da Acetil-CoA Carboxilase, funciona principalmente como inibidor da oxidação de ácidos graxos
Citrato alostericamente ativa Acetil-CoA Carboxilase. [Citrato] é alta quando há quantidade adequada de acetil-CoA entrando no ciclo dr Krebs. Excesso de acetil-CoA é então convertido via malonil-CoA em AG para estoque
Síntese de ácido graxo ocorre devido a uma série de reações: • Em mamíferos é catalizada por 7 domínios individuais de único polipeptídio. • NADPH serve como doador deelétrons em duasreações envolvendo redução de substrato. NADPH é produzido principalmente nas via das Pentoses Fosfato.
A reacao de condensacao (passo 3) envolve descarboxilacao do malonil seguido do ataque do carbanion resultante no carbono carbonil do acetil (ou acil).
A b-cetona e reduzida a um alcool pela transferencia de e- do NADPH. • Desidratacao rende dupla em trans. • Reducao pelo NADPH rende cadeia saturada”.
Segue-se transferencia do AG crescente da fosfopanteteina para a enzima de Condensacao (residuo sulfifril da cisteine) e o ciclo comeca de novo com a entrada de u lutro malonil.
Liberação do Produto Quando o AG tem 16 carbonos, o domínio da Tioesterase catalisa a hidrolise do tioéster que liga o AG à fosfpanteteina. O AG de 16-C saturadopalmitato é o produto final do complexo AG sintase.
AG Sintase é um dímero. Em cada monômero, a ordem das enzimas é a mostrada abaixo. Os 2 monômeros são antiparalelos. Cada passo de transferência ocorre da enzima de condensação de um lado para ACP do outro .
A transcrição da AG Sintase is regulada No fígado: • Insulina, estimula a expressão da AG Sintase. Assim, o excesso de glicose é estocado como gordura. • Fatores de transcrição medeiam o efeito estimulatório da insulina, incluindo USFs (upstream stimulatory factors) and SREBP-1. • SREBPs (sterol response element binding proteins) foram primeiro identificadas pelo seu papel na regulação da síntese de colesterol. • AG Poliinsaturados diminuem atranscrição do gene da AG Sintase no hepatócito por suprimir SREBPs.
A transcrição da AG Sintase is regulada No Adipócito: • Expressão de SREBP-1 e AG Sintase é inibida por leptina, um hormonio que tem papel na regulação da ingestão alimentar e metabolismo de gordura. • Leptina é produzida por adipócitos em resposta ao excesso de estoque de gordura. • Leptina regula peso corporal por diminuir ingestãi alimentar, aumentando dispêndio de energia e inibindo a síntese de AG.
Elongação além de 16-C ocorre na mitocôndria and reticulo endoplasmico (ER). • mitocôndria involve b-oxidation no reverso (ver adiante), exceto que NADPH serve como doador eletron. • Poliinsaturados esterificados com coA são substratos para ER elongação,que usa malonyl-CoA como doador de 2 carbonos.
Desaturases colocam duplas em posições especificas na cadeia. Mamíferos incapazes de produzir duplas em alguns locais,p.ex D12. Assim, AG PI é alguns AG são essenciais como ac linoléico 18:2 cis D9,12.
Dessaturação envolve proteínas do RE : • NADH-cyt b5 Redutase, flavoproteina com FAD como grupo prostético. • Citochrome b5, proteína separada ou domínio no final da dessaturase. • Dessaturase, com o sítio ativo com 2 átomos de ferro complexados em resíduos de histidina.
Dessaturase são oxidase de função mista. • Redução de 4e- do O2 2 H2O para formar a dupla. • Os e- passam do NADH para a dessaturase via redutase contendo FAD cytochrome b5, NADH FAD cyt b5 desaturase • 2e- são extraídos de AG para formar a dupla. • estearato + NADH + H+ + O2oleato + NAD+ + 2H2O
DEGRADAÇÃO DE TRIACILGLICEROL: Epinefrina, norepinefrina, glucagon, ACTH todos ativam a lipase sensível a hormônio via Adenilato ciclase
Glicerol fígado – oxidado a diidroxiacetona 3 fosfato (glicólise ou neoglicogênese) • AG vão para o sangue ligados à albumina • AG entra na célula e liga-se a CoA (acil CoA sintetase) • AG + ATP = Acil-AMP + PPi (PPi = 2Pi pela pirofosfatase) • Acil-AMP + CoASH = AcilCoA + AMP
Outra volta Atua normalmente AG inaturados: 2 enzimas : ISOMERASE E REDUTASE. Isomerase transforma cis D3 em trans D2.. Daqui para frente normal. Cis D3 enoil CoA não é substrato para a desidrogenase. (Dupla C3-4 impede a C2-C3) Cria Cis D3 enoil CoA Isomerase transforma cis D3 em trans D2.. Daqui para frente normal. Cria intermediário que não é substrato pra hidratase
CORPOS CETÔNICOS TIOLASE