Download
1 / 39
1.15k likes | 4.28k Views
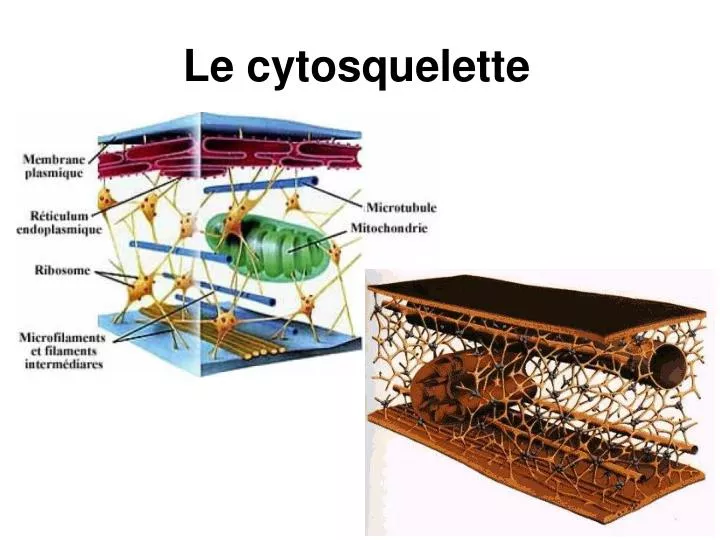
Le cytosquelette. Le cytosquelette. Reseau formee de trois types de proteines filamenteuses Filaments d’actine Microtubules Filaments intermediaires. Le cytosquelette. DIMENSIONS (DIAMETRES) les microfilaments (5-9nm - filaments d'actine) les filaments intermédiaires (10nm)

E N D
Le cytosquelette Reseau formee de trois types de proteines filamenteuses Filaments d’actine Microtubules Filaments intermediaires
Le cytosquelette DIMENSIONS (DIAMETRES) les microfilaments (5-9nm - filaments d'actine) les filaments intermédiaires (10nm) les microtubules (25nm)
Le cytosquelette POSITION DES ELEMENTS DANS LA CELLULE
MICROFILAMENTS D’ACTINE 7 types moleculaires d’actine: 4 isoformes de -actine muscles striés squelettiques et cardiaque et les muscles lisses) 2 isoformes de -actine muscle lisse entérique et dans les tissus non musculaires 1 type de -actine, m. lisse intestinal non musculaire 2 formes d’organisation: actine G – forme globulaire actine F – forme fibrilaire
MICROFILAMENTS D’ACTINE Actine globulaire - 1 polypeptide PM 45000 Da, 6 nm / 4 nm. sites de couplage pour: profiline (protéine inhibitrice pour la polymérisation), pour Ca, Mg – contrôle de la polymérisation, pour ATP (énergie pour la polymérisation) myosine (tete).
MICROFILAMENTS D’ACTINE Actine fibrillaire par polymérisation de l’actine G, demande ATP, Ca, Mg
MICROFILAMENTS D’ACTINE L’actine G se polymérise en présence d’ATP (lié à l’actine) et le filament formé est une structure polaire. Il a une extrémité à croissance rapide (nommée « + ») puisque environ 1000 monomères peuvent être ajoutés par seconde et une extrémité à croissance lente, voire très lente, (nommée « - » ). De nombreuses protéines (ARP2/3= Actin Related Proteins ; CapZ) initient la polymérisation alors que la gelsoline peut créer une scission du filament et éviter la repolymérisation.
Les propriétés contractiles des filaments d'actine sont dûes à une famille de protéines motrices spécifiques : les myosines. LES MYOSINES proteine abondente dans l’organisme (7 -14 classes) deux chaînes lourdes (dimère) de 2 000 acides aminés dont les extrémités C-terminales en hélice α sont surenroulées, ce qui confère à la myosine la forme rigide d'un bâtonnet de 180 µm, et dont les têtes N-terminales de 190 acides aminés constituent les domaines moteurs. deux paires de chaînes légères : une paire de chaînes légères dites essentielles (ELC), et une paire de chaînes légères dites régulatrices (RLC) qui stabilisent la longue hélice près du domaine N-terminal (tête), dans la région du cou qui forme le ‘bras de levier’.
La myosine=protéine qui joue un rôle fondamental dans les mécanismes de la contraction musculaire. Elle se rencontre donc dans les cellules à activité contractile des vertébrés, telles les cellules musculaires. LES MYOSINES La myosine II est une molécule allongée de 2x240 kDa composée de deux chaînes lourdes (environ 200 kDa chacune) et de quatres chaînes légères (environ 20 kDa chacune).
Chaque chaîne lourde est constituée par • une queue C-terminale allongée et fibrillaire en hélice alpha, • une tête globulaire N-terminale enzymatique à activité ATPasique associée à • deux chaînes légères et • un domaine cervical déformable reliant les deux extrémités. • Tête globulaire et partie cervicale forment la méromyosine lourde, • La partie fibrillaire caudale formant la méromyosine légère. • Les queues allongées de deux chaînes lourdes de myosine s'enroulent l'une autour de l'autre en une superhélice, les deux têtes globulaires se trouvant côte à côte LES MYOSINES
Contraction – regulee par 4 proteines accessoires: • Tropomyosine • 3 troponines, type C, I, T. • Tropomyosine (Tm) • - 40 nm; • - Chaine continue a travers le filament d’actine • - Chaque Tm - 7 sites pour l’actine. • Troponines • - la troponine-T est responsable de la liaison troponine-tropomyosine; • la troponine-I possède une activité inhibitrice de l'activité ATPasique de la myosine; • la troponine-C possède 4 sites de fixation pour le calcium qui, lorsqu'ils sont occupés, lèvent l'action de la troponine I INTERACTION ACTINE-MYOSINE
INTERACTION ACTINE-MYOSINE Plusieurs centaines de molécules de myosines II s'assemblent pour former un filament épais. Les parties caudales de ces molécules sont rassemblées parallèlement. Les têtes globulaires dépassent en périphérie de ce filament et sont donc disponibles pour pouvoir se fixer aux filaments d'actine. Les molécules de myosine étant disposées en deux groupes tête-bèche, la partie centrale du filament (correspondant à la strie M) est dénudée, c'est à dire dépourvue de tête globulaire
INTERACTION ACTINE-MYOSINE • Phase 0 • Situs de couplage ATP – nu • Tete de myosine couple a l’actine • Phase 1 • ATP couple la myosine • Tete myosine – enlevée • Hydrolyse ATP- ADP+Pi • Site ATP – fermeture • Pliage tete myosine sur le prochain monomere d’actine • Phase 2 • Couplage myosine –prochaine actine • Delivrer Pi, ADP • Traction filament d’actine • Phase 3 • Delivrer ADP • Revenir au position initiale
Les microtubules sont présents dans l’ensemble des cellules eucaryotes. Ils peuvent être fortement représentés dans certains types cellulaires comme les neurones où ils ont un rôle fondamental dans - le transport axonal des vésicules, - ou dans des cellules à forte activité sécrétoire MICROTUBULES Constitution : Ils sont constitués de dimères de tubuline α et β. Chaque monomère peut s’associer au GTP mais seule la tubuline β peut échanger son GTP. C’est une protéine globulaire comme l’actine
Assemblage : Microtubules = structures polaires avec - une extrémité (+) à croissance rapide dirigée vers la périphérie de la cellule et - une extrémité (-) qui est associée au centrosome. Le centrosome est un complexe protéique situé près du noyau et il est constitué de deux centrioles eux-mêmes constitués de tubuline α, β, γ, δ et ε. MICROTUBULES
Assemblage - etapes: • polymérisation de dimères de tubuline a et b (chargées de GTP). Les dimères s’associent tête bêche pour former un protofilament. Après polymérisation le GTP de la tubuline b est hydrolysé en GDP. • formation d’un fragment de microtubule par association latérale de 10 à 15 protofilaments et repliement du feuillet pour donner une structure rigide. • élongation du microtubule par polymérisation (ajout de dimères) à l’extrémité (+) MICROTUBULES
Centres organisateurs des microtubules (MTOC).(centrosomes) • réseau radiaire perinucleaire pour organiser les microtubules • au niveau du MTOC, les microtubules presentent l’extremite (-), pour croissance radiaire. • MTOC comprend une paire de centrioles.
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
FACTEURS STABILISATEURS ET DESTABILISATEURS DES MICROTUBULES
Diamètre (10 nm) intermédiaire entre les microfilaments d’actine (7 nm) et les microtubules (24 nm). • FI extrêmement stables • Structures fibrillaires -héliçages s’assemblant en des filaments • FI n’ont las la capacité de coupler des nucléotides FILAMENTS INTERMEDIAIRES