Download

1 / 41
410 likes | 572 Views
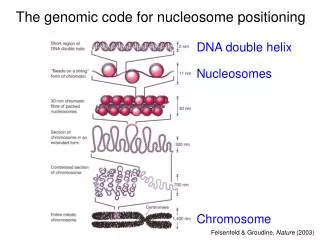
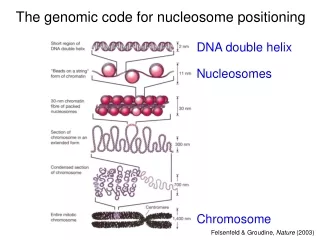
A genomic code for nucleosome positioning. Hierarchical DNA folding in eukaryotic chromosomes. DNA double helix. Nucleosomes. Felsenfeld & Groudine,, Nature 421: 448-453 (2003). Most eukaryotic DNA is sharply looped. Luger et al., 1997. Random 220-mer pool

E N D
Hierarchical DNA folding in eukaryotic chromosomes DNA double helix Nucleosomes Felsenfeld & Groudine,, Nature 421: 448-453 (2003)
Most eukaryotic DNA is sharply looped Luger et al., 1997
Random 220-mer pool (1 each of 5 x 1012 individuals) PCR amplify, HPLC purify Select best 10% (dialysis from 2.0M NaCl) 2x Sucrose gradient purification Extract DNA Population diversity analysis Free energy analysis Clone, sequence, analyze individuals Physical selection for stable nucleosome formation on chemically synthetic random DNAs Lowary & Widom, 1998
Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation Lowary & Widom, 1998 Thåström et al., 1999 Widom, 2001 Thåström et al., 2004
Basepair steps as fundamental units of DNA mechanics Zhurkin Olson
AATTTA GC AATTTA GC AATTTA GC GC AATTTA AATTTA GC GC AATTTA AATTTA GC DNA sequence motifs that stabilize nucleosomes and facilitate spontaneous sharp looping Segal et al., 2006
Why DNA some sequences have especially high affinity for histone octamer • More or better bonds • Appropriately bent • More easily bendable • Appropriate twist • More easily twistable Widom, 2001 Cloutier & Widom, 2004
~10.2 bp periodicity of AA/TT stepsin the C. elegans genome Widom, 1996
Collect nucleosome-bound sequences Validate model Construct nucleosome-DNA interaction model Compare within vivo positions Predict intrinsically encodednucleosome organization Associate intrinsic encoding with biological function Understanding and predicting the genome’s nucleosome-forming potential Segal et al., 2006
Isolation of natural nucleosome core DNA Alberts et al., 4th ed., Fig. 4–24 (2002)
ACGTAGCTGTAGTGTACTGACGTACGTCGTC ACTAGCTGATACGGAGACCCGCGCGATTTTGCGGTC ACTGTTCGTCGTGTGTGTGTGCTGCTGTAGACTTGTGTG TACTGTTTTTATTTTGCGGGCATGCTTGT Center align ACGTAGCTGTAGTGTACTGACGTACGTCGTC ACTAGCTGATACGGAGACCCGCGCGATTTTGCGGTC ACTGTTCGTCGTGTGTGTGTGCTGCTGTAGACTTGTGTG TACTGTTTTTATTTTGCGGGCATGCTTGT Center alignment of yeast nucleosome DNAs • ~10bp periodicity of AA/TT/TA • Same period for GC, out of phase with AA/TT/TA • Signals important for DNA bending • NO signal from alignment of randomly chosen genomic regions Yeast (in vivo nucleosomes) Segal et al., 2006
Center alignment Location mixture model alignment Location mixture model alignment vs center alignment Wang & Widom, 2005
Center alignment of chicken nucleosome DNAs Chicken + Yeast merge Chicken (in vivo) (Satchwell et al., 1986) Yeast (in vivo) Segal et al., 2006
Alignments of nucleosomes selected in vitro Mouse (in vitro) (Widlund et al., 1997) Random DNA (in vitro)(Lowary & Widom, 1998) Yeast (in vitro) Chicken (in vivo) (Satchwell et al., 1986) Yeast (in vivo) Segal et al., 2006
In vitro experimental validation of histone-DNA interaction model • Adding key motifs increases nucleosome affinity • Deleting motifs or disrupting their spacing decreases affinity Segal et al., 2006
In vitro experimental validation of histone-DNA interaction model • Adding key motifs increases nucleosome affinity • Deleting motifs or disrupting their spacing decreases affinity dyad 38 8 18 28 48 58 68 78 88 98 108 118 128 138 Segal et al., 2006
Summary Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation We have a predictive understanding of the DNA sequence motifs that are responsible
AATTTA GC AATTTA GC AATTTA GC GC AATTTA AATTTA GC GC AATTTA AATTTA GC Nucleosome–DNA model Differences from TF model • Signal is weak • Information in dinucleotides • Two-fold symmetry axis • Position specific dinucleotide distributions • Add reverse complement of each sequence • Local (3 bp) averaging Segal et al., 2006
Summary Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation We have a predictive understanding of the DNA sequence motifs that are responsible Sequences matching these motifs are abundant in eukaryotic genomes, and are occupied by nucleosomes in vivo
Log likelihood Genomic Location (bp) Placing nucleosomes on the genome A free energy landscape, not just scores and a threshold !! • Nucleosomes occupy 147 bp and exclude 157 bp Segal et al., 2006
Equilibrium configurations of nucleosomeson the genome • One of very many possible configurations P(S) PB(S) P(S) PB(S) P(S) PB(S) P(S) PB(S) Chemical potential – apparent concentration Probability of placing a nucleosome starting at each allowed basepair i of S Probability of any nucleosome covering position i ( average occupancy) Locations i with high probability for starting a nucleosome ( stable nucleosomes) Segal et al., 2006
Cross-correlation of average occupancies predicted using yeast and chicken models Segal et al., 2006
Nucleosome coding potential at the GAL1–10 locus: predicted distribution compared to experimental Segal et al., 2006
Nucleosome coding potential at the CHA1 locus Segal et al., 2006
Yeast intergenics depleted of nucleosomes (Bernstein et al.) Yeast intergenics occupied by nucleosomes (Bernstein et al.) Reliable classification of nucleosome occupancies on depleted vs occupied regions Yeast ORFs depleted of nucleosomes (Lee et al.) (0.82, 0.68) Yeast ORFs occupied by nucleosomes (Lee et al.) (0.88, 0.76) (0.88, 0.55) p<10–6 p<10–9 (0.82, 0.32) Segal et al., 2006
High Predictions + Controls – Controls In vivo nucleosome occupancies at predicted high-occupancy locations Segal et al., 2006
In vivo nucleosome occupancies at predicted low-occupancy locations Segal et al., 2006
Summary Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation We have a predictive understanding of the DNA sequence motifs that are responsible Sequences matching these motifs are abundant in eukaryotic genomes, and are occupied by nucleosomes in vivo A model based only on these DNA sequence motifs and nucleosome-nucleosome exclusion explains ~50% of in vivo nucleosome positions
Understanding and predicting the genome’s nucleosome-forming potential Collect nucleosome-bound sequences Validate model Construct nucleosome-DNA interaction model Compare within vivo positions Predict intrinsically encodednucleosome organization Associate intrinsic encoding with biological function Segal et al., 2006
Average nucleosome occupancy tRNAs Telomeres Centromere Ribosomal RNAs Divergent intergenic regions Conserved DNA binding sites Unique intergenic regions Convergent intergenic regions Ribosomal proteins Autonomously replicating sequences Protein coding regions Nucleosome occupancy varies with chromosome region type Segal et al., 2006
Does the genome’s intrinsic nucleosome organization facilitate occupancy of functional binding sites? Segal et al., 2006
The yeast genome encodes low nucleosome occupancy over functional binding sites Segal et al., 2006
Model Permuted Semi-stable nucleosomes Semi-stable nucleosomes Stable nucleosome TATA Box Distinctive nucleosome occupancy adjacent to TATA elements at yeast promoters Segal et al., 2006
-36 -36 S. cerevisiae -41 S. Paradoxus -22 S. mikatae -41 S. kudriavzevii -42 S. bayanus -23 S. castelli -42 C. glabrata -21 S. kluyveri -16 K. walti -53 K. lactis -14 -9 A. gossypii -26 D. hansenii -56 C. albicans Y. lipolytica -3 A. nidulans -2 S. pombe -3 Nucleosome organization near 5’ ends of genes is conserved through evolution Segal et al., 2006
Predicted nucleosome organization near 5’ ends of genes – comparison to experiment Segal et al., 2006
Summary Differing DNA sequences exhibit a > 5,000-fold range of affinities for nucleosome formation We have a predictive understanding of the DNA sequence motifs that are responsible Sequences matching these motifs are abundant in eukaryotic genomes, and are occupied by nucleosomes in vivo A model based only on these DNA sequence motifs and nucleosome-nucleosome exclusion explains ~50% of in vivo nucleosome positions These intrinsically encoded nucleosome positions are correlated with, and may facilitate, essential aspects of chromosome structure and function
Promoter region 1 Promoter region 2 0 100 200 0 100 200 20 0 -20 I None 1 0 II Factors present in the nucleus 1 0 III + 1 0 / Nucleosome score landscape Stable nucleosome DNA binding protein / Nucleosome occupancy Unstable nucleosome DNA binding site Genome sequence influences the competition between nucleosomes and DNA binding proteins Segal et al., 2006
AATTTA GC AATTTA GC AATTTA GC GC AATTTA AATTTA GC GC AATTTA AATTTA GC Toward a proper free energy model for the sequence-dependent cost of DNA wrapping Morozov, Segal, Widom, & Siggia
Acknowledgements Nucleosome positioning in vivo Eran Segal (Weizmann Inst.) Yvonne Fondufe-Mittendorf Lingyi Chen Annchristine Thåström Yair Field (Weizmann Inst.) Irene Moore Jiping Wang (Northwestern U. Statistics) Alexandre Morozov (Rockefeller U.) Eric Siggia (Rockefeller U.)